http://www.youtube.com/watch?v=T_f_fGUpwJU&feature=player_embedded
aqui esta nuestro video del olfato Gabriela García Salazar 210352702
martes, 17 de julio de 2012
domingo, 15 de julio de 2012
Transducción olfativa
http://www.youtube.com/watch?v=PwYFGua19T4
El sentido del olfato, al igual que el sentido del gusto, es un sentido químico. Se denominan sentidos químicos porque detectan compuestos químicos en el ambiente, con la diferencia de que el sentido del olfato funciona a distancias mucho más largas que el sentido del gusto. El proceso del olfato sigue más o menos estos pasos:
1.Las moléculas del olor en forma de vapor (compuestos químicos) que están flotando en el aire llegan a las fosas nasales y se disuelven en las mucosidades (que se ubican en la parte superior de cada fosa nasal).
2.Debajo de las mucosidades, en el epitelio olfatorio, las células receptoras especializadas, también llamadas neuronas receptoras del olfato, detectan los olores. Estas neuronas son capaces de detectar miles de olores diferentes.
3.Las neuronas receptoras del olfato transmiten la información a los bulbos olfatorios, que se encuentran en la parte de atrás de la nariz.
4.Los bulbos olfatorios tienen receptores sensoriales que en realidad son parte del cerebro que envían mensajes directamente a:
◦los centros más primitivos del cerebro donde se estimulan las emociones y memorias (estructuras del sistema límbico) y
◦centros “avanzados” donde se modifican los pensamientos concientes (neocorteza).
5.Estos centros cerebrales perciben olores y tienen acceso a recuerdos que nos traen a la memoria personas, lugares o situaciones relacionadas con estas sensaciones olfativas.
El sentido del olfato, al igual que el sentido del gusto, es un sentido químico. Se denominan sentidos químicos porque detectan compuestos químicos en el ambiente, con la diferencia de que el sentido del olfato funciona a distancias mucho más largas que el sentido del gusto. El proceso del olfato sigue más o menos estos pasos:
1.Las moléculas del olor en forma de vapor (compuestos químicos) que están flotando en el aire llegan a las fosas nasales y se disuelven en las mucosidades (que se ubican en la parte superior de cada fosa nasal).
2.Debajo de las mucosidades, en el epitelio olfatorio, las células receptoras especializadas, también llamadas neuronas receptoras del olfato, detectan los olores. Estas neuronas son capaces de detectar miles de olores diferentes.
3.Las neuronas receptoras del olfato transmiten la información a los bulbos olfatorios, que se encuentran en la parte de atrás de la nariz.
4.Los bulbos olfatorios tienen receptores sensoriales que en realidad son parte del cerebro que envían mensajes directamente a:
◦los centros más primitivos del cerebro donde se estimulan las emociones y memorias (estructuras del sistema límbico) y
◦centros “avanzados” donde se modifican los pensamientos concientes (neocorteza).
5.Estos centros cerebrales perciben olores y tienen acceso a recuerdos que nos traen a la memoria personas, lugares o situaciones relacionadas con estas sensaciones olfativas.
Química de los olores
http://www.youtube.com/watch?v=2jyXUrgkexo
¿Qué es el olor?
Desde el punto de vista químico, el olor es una sensación, una noción de estímulo y percepción producida en el olfato por la interacción de una sustancia orgánica con los receptores olfativos de los seres vivientes. Dicha interacción depende en gran medida de la volatilidad de la sustancia, ya que es necesario que las moléculas de la sustancia olorosa pasen a una fase gaseosa para que puedan llegar a la nariz y así ser percibidos; de igual forma, se requiere que puedan atravesar las membranas de las células epiteliales de la nariz y llegar a los receptores que enviarán la señal al cerebro, indicando la sensación del olor. Un punto importante que deben cumplir las moléculas olorosas es tener un peso molecular bajo, aunque se ha generado cierta controversia en este punto puesto que hay moléculas, tales como los esteroides, que poseen olor aunsiendo de gran tamaño. Otro aspecto en relación con el tamaño de la molécula es su influencia sobre el mecanismo de la quimiorrecepción, pues parece ser que cier tas incapacidades para percibir aromas (anosmias) en los humanos aumentan directamente conforme al tamaño molecular, lo que puede deberse a que las moléculas muy grandes no tienen un buen acomodo en el sitio receptor.
¿Cómo percibimos los olores?
La percepción del olfato involucra tres aspectos importantes: la intensidad, la descripción cualitativa y la apreciación del aroma. La relación entre la concentración de un olor y la intensidad percibida es bien conocida y sigue una distribución logarítmica común, al igual que la de otros sistemas sensoriales. El umbral de detección del olor, definido como la concentración mínima en que se percibe el estímulo, puede ser hasta de partes por trillón para algunos aromas, pero esta estimación varía de un aroma a otro hasta en unas cincuenta veces o más.
Cuando la descripción cualitativa se compara con la intensidad, se presenta una ligera complicación; se cree que los humanos tienen la capacidad de diferenciar hasta 10 mil olores diferentes, pero la mayoría de nosotros solamente percibimos una fracción de ellos a lo largo de la vida, por lo que a menudo la descripción de un olor varía de persona a persona.
En la actualidad se conocen miles de olores; sin embargo, la apreciación de un aroma es subjetiva, ya que se ve influenciada por aspectos culturales o emocionales particulares, es decir, de cada sujeto; de hecho, para algunas personas el mismo perfume puede ser agradable, desagradable o indiferente. No obstante, los estudios fisiológicos enfocados a esta área, basados en pruebas psicológicas, sostienen que muchas sensaciones olfatorias dependen de unas cuantas emociones primarias. Por ello –independientemente de la forma particular en la que cada individuo percibe los olores–, se ha propuesto una sencilla clasificación que postula siete aromas primarios: etéreo, alcanforado, almizcle, floral, mentolado, picante y pútrido. Aunque ésta ha sido la tipificación usualmente aceptada por los científicos, hasta ahora no hay un “mapa de olores” que se ajuste a todos los individuos y que se utilice por las industrias de la cosmética, perfumería y alimentaria. Además, algunos reportes indican que las sensaciones primarias pueden ser hasta cincuenta respuestas o más; sin embargo, es probable que a pesar de que esa cantidad sea significativa, las que puede experimentar el ser humano son solamente las propuestas arriba, argumentando que se posee una cierta “ceguera” para el olor o una ausencia de receptores adecuados para la identificación y caracterización de la molécula que causa tal sensación.
El sentido del olfato
El sentido del olfato es una de las ventanas más sofisticadas a través de la cual un individuo capta información del ambiente externo y por la cual los seres vivos de cualquier especie son capaces de modificar su comportamiento y establecer su gusto por determinados ambientes y alimentos.
En el caso de los humanos, el aparato olfativo tiene varias diferencias respecto del que poseen animales e insectos; incluso se puede decir que nuestro sentido del olfato es extremadamente rudimentario comparado con el de otras especies.
Probablemente esta curiosa diferencia se deba a que la especie humana ha alcanzado a lo largo de su evolución un desarrollo más acentuado de los demás sentidos y también de otras aptitudes, relegando en cierta forma la capacidad olfativa, dando razón de por qué nosotros tenemos otras forma de comunicación distintas de un “simple” sendero aromático.
Paradójicamente, el sentido del olfato de todos los mamíferos conserva ciertas similitudes en la cavidad nasal, tales como su localización y asociación con las vías respiratorias superiores, puesto que los compuestos olorosos se hallan dispersos en el aire, y es así como logran llegar a la nariz para alcanzar las células olfatorias y estimularlas.
Por su parte, la cavidad nasal se encuentra protegida en su totalidad por una capa de células epiteliales y, curiosamente, sólo alrededor de 5% del epitelio que la cubre está relacionada con la función de detectar e identificar los aromas. El epitelio olfativo mide 2.5 cm2 y contiene aproximadamente cincuenta millones de células, mismas que albergan los receptores químicos responsables de la identificación de los olores; dichas células son estructuras provenientes del sistema nervioso central.
Resulta interesante que el epitelio olfativo esté constituido de tres capas principales de células: las células de soporte, que tienen la función de aislar eléctricamente las neuronas; las células receptoras del olfato, que son las responsables de la transmisión del estímulo, y las células basales, que se encuentran en la parte más recóndita del epitelio. Se puede decir entonces que el sentido del olfato no es más que una serie de experiencias dependientes del funcionamiento cerebral y que opera siempre mediante mensajes neuronales.
Algunos científicos consideran que el olor es una percepción que no puede ser medida, en la que el sistema límbico (emocional) es el que afecta la apreciación para clasificar el agrado o desagrado por los aromas, además de que es inf luido por algunos aspectos de orden f isiológico. Tomemos como ejemplo la comida. Cuando se está cocinando o con hambre, el olor tiene una percepción grata; sin embargo, se vuelve desagradable cuando ya se ha comido.
El mecanismo bioquímico del sentido del olfato
A diferencia de la vista, que se explica en función de las leyes fundamentales de la óptica, o de la audición, entendida por fenómenos armónicos, el sentido del olfato tiene aún ciertas incógnitas. La percepción de los aromas, así como el sistema olfativo en general, es de los mecanismos biológicos menos estudiados, probablemente por su localización y, por ende, por su dificultad para reproducirse en un laboratorio. Pese a ello, en el campo de la bioquímica se especula que la generación del impulso nervioso como respuesta a una molécula capaz de producir un aroma es el resultado de una variación de voltaje en la membrana celular, originado por una alteración en los niveles de monofosfato de adenosina cíclico o inositol trifosfato, proceso químico y eléctrico bien conocido que permite postular tres mecanismos hipotéticos mediante los cuales se puede llevar a cabo la traducción del mensaje olfativo: a) por la apertura directa de los canales iónicos alcontacto con la molécula del olor, b) por la alteración de la fluidez de la membrana al interactuar los receptores con los aromas, y c) por una transmisión indirecta del olor, a través de proteínas receptoras ligadas a rutas, dependiente de segundos mensajeros.
Algunos estudios recientes han revelado que el código de olores es diferenciado por múltiples receptores, y que diferentes olores se reconocen por sus distintas combinaciones, de modo que la selección no es específica, sino que se sitúa en un nivel molecular; a tal selección se le ha denominado reconocimiento odotópico. Tal hipótesis ha sido ampliamente aceptada por dos sencillas razones: la primera es que no podrían existir millones de proteínas receptoras, una para cada olor, y la segunda se basa en estudios electrofisiológicos en los que se ha encontrado que una neurona olfativa responde a otras moléculas olorosas de estructura química diferente.
Adicionalmente se ha sugerido que existe agonismo o antagonismo entre los olores y sus receptores en el epitelio olfativo; así, hay una competencia entre las combinaciones de aromas frente a los agonistas para activar un receptor neuronal, o entre agonistas y antagonistas para bloquear una neurona. La incapacidad de un agonista para fijarse en un sitio receptor y activar lo consti tuye uno de los mecanismos más efectivos para provocar una disminución en la percepción del olor; dicha reducción puede considerarse como supresión del olor, y es lo que comúnmente se pretende hacer mediante cier tas formulaciones que
enmascaran los sabores y olores desagradables, como ciertos alimentos, medicinas, cosméticos o productos de limpieza.
Teorías del olfato
Se han propuesto diversas teorías para comprender el funcionamiento biológico de diferenciación del sentido del olfato, tratando de dar una explicación coherente y plausible del modo en que interactúan los olores y los receptores olfativos; sin embargo, debido a la carencia de conocimiento en este campo, muchas de ellas han fracasado o no han sido adecuadamente demostradas. La teoría más aceptada para tal fin es la teoría estereoquímica, que involucra la relación de la estructura tridimensional de la molécula con las propiedades olfativas de los compuestos. Esta teoría se basa en la idea de que los grupos funcionales presentes en una molécula odorífica tienen distintos tamaños, y que mediante un mecanismo similar al modelo enzimático “llave-cerradura”, se puede reconocer la distribución electrónica de un compuesto asociado a un olor característico, siempre y cuando ambas partes coincidan con el arreglo conformacional y una distribución espacial específica.
Olor y estructura molecular
En el ámbito de la investigación farmacéutica, cosmética y de al imentos, la int roducción de nuevos compuestos con propiedades biológicas activas de interés implica un alto costo, ya que requiere técnicas y métodos de laboratorio que, además, necesitan procesos de control y regulación. Por ello se ha venido desarrollando recientemente una metodología para el estudio de nuevos compuestos con cualidades incluso personalizadas, pero que evita una gran inversión económica, ya que el objetivo se logra a través de métodos computacionales que permiten correlacionar la estructura química con la actividad biológica. Estas técnicas hacen posible un análisis adecuado de la estructura molecular, bajo el precepto de que tal relación implica la pertenencia al mismo tipo de estructura y, por consiguiente, a la misma clase de aroma. Así, el diseño computacional de fragancias reúne en el campo de la investigación las bases de la ciencia, la experiencia y el instinto, que se conjugan como si se tratara de una fantasía creacional. A la fecha hay varias reglas para crear agentes odoríficos basados en la estructura química de las sustancias. A continuación se presentan unos cuantos hallazgos de cómo la estructura puede emplearse para predecir o simplemente especular acerca del aroma que un cierto compuesto puede provocar.
Ambergris
Es una fragancia de origen animal secretada en el estómago o tracto intestinal de la ballena y que se libera al mar en forma de una masa rocosa de color grisáceo a negro. Cuando se expone a la luz, al aire y al agua marina, el material se degrada a un color amarillo cremoso, al mismo tiempo que se oxida su principal componente, el alcohol triterpénico ambreina. De acuerdo con el análisis estructural de las propiedades olfativas se ha logrado establecer que debe haber un anillo de trans-decalina dentro de la molécula para dar esta clase de aroma.
Almendras amargas
Este grupo de olores ha sido el modelo por excelencia en los estudios de la relación estructura-actividad, puesto que tiene un olor bien definido. Las moléculas clásicas poseedoras de dicho aroma son el ácido cianhídrico y el benzaldehído, ambos producidos por la hidrólisis de la amigdalina, que es uno de los componentes de las almendras amargas. Sin embargo, si se compara la estructura de ambos compuestos se puede deducir una gran variabilidad estructural, por lo que después de ciertos estudios computacionales se ha logrado establecer que este aroma debe tener un núcleo aromático; cuando los dobles enlaces se encuentran conjugados con grupos funcionales, el aroma se intensifica.
Alcanforado
Los aromas alcanforados se han popularizado por su presencia en productos tales como los ungüentos para tratar resfriados y en antiguas formulaciones de repelentes de polillas. La molécula líder responsable de este olor es el alcanfor, que se puede obtener de forma natural a través del aceite de madera o en el laboratorio, a partir del pineno.
Floral
Las fragancias florales son de las más interesantes dada su gran aceptación por parte de la mayoría de las personas, sobre todo de las mujeres, quienes dominan la industria del perfume. Se pueden usar cientos de diferentes notas florales, siendo las más importantes la rosa, el jazmín y el lirio de los valles. De estos tres solamente se han realizando análisis de estructuraactividad con los dos últimos, puesto que el perfume de rosas es un olor complejo. No obstante, se ha logrado revelar ciertas características moleculares para los olores florales; por ejemplo, sustituyendo anillos de tamaño mediano con grupos funcionales del tipo del alcohol, aldehído, éster o éter.
Frutal
De las fragancias frutales más utilizadas los ésteres forman parte del grupo de compuestos que más se ha estudiado. Se sabe que la intensidad del olor depende considerablemente de la naturaleza de la porción ácida y alcohólica; para los ésteres alifáticos se ha propuesto que aquellos con más de ocho átomos de carbono tienen aromas frutales intensos, y adicionalmente se ha sugerido que las insaturaciones en la proximidad de la función éster reducen el peculiar tono frutal y agregan sensaciones herbales. Por su parte, los olores a uva, frutas rojas, melón y otros de percepción similar se producen cuando ciertos compuestos azufrados tienen bajas concentraciones; de hecho, muchos compuestos orgánicos volátiles de este tipo se utilizan enormemente en la industria de bebidas y alimentos, y no simplemente para productos derivados de frutas, sino para la carne, pan, papas, cerveza y café.
Herbal
Esta vasta familia de fragancias ha sido objeto de muy pocos estudios que relacionen la estructura con el aroma, probablemente porque este aroma es muy difícil de definir. La nota herbal alude tanto al follaje verde de las hojas, tallos y frutos verdes, como al pasto recién cortado. La principal sustancia vinculada con este olor es el cis-3-hexen-1-ol, muchos de cuyos derivados se han introducido en la industria de los sabores y fragancias desde 1960.
Musk
Los aromas del tipo musk son de los más interesantes en el ámbito de la perfumería por dos razones: su gran capacidad como
fijador y sus propiedades características. Son olores considerados cálidos, sensuales y a la vez naturales. Este aroma está químicamente bien definido y abarca cuatro diferentes grupos: macrocíclos nitrogenados, no nitrogenados, bencenoides y esteroidales.
Maderas
En este grupo se incluyen las fragancias derivadas de los aceites esenciales de varios tipos de árboles, como el cedro y el sándalo, y algunos provenientes de hojas como el pachuli y el vetiver. Generalmente son compuestos bicíclicos o tricíclicos de 12 a 17 carbonos y con un grupo funcional alcohol o éster.
El desarrollo e innovación de las fragancias puede en algunos casos considerarse como un capricho meramente placentero, sin embargo, el estudio racional de la relación estructuraactividad de los olores ha permitido comprender algunas semejanzas estructurales en las moléculas de los aromas.
Quizás el enigma del olor entraña, más allá de la fisiología cognitiva, ciertos procesos de comunicación, comportamientos antropológicos y, sobre todo, un mundo de comunicación química que aún está por descubrirse.
¿Qué es el olor?
Desde el punto de vista químico, el olor es una sensación, una noción de estímulo y percepción producida en el olfato por la interacción de una sustancia orgánica con los receptores olfativos de los seres vivientes. Dicha interacción depende en gran medida de la volatilidad de la sustancia, ya que es necesario que las moléculas de la sustancia olorosa pasen a una fase gaseosa para que puedan llegar a la nariz y así ser percibidos; de igual forma, se requiere que puedan atravesar las membranas de las células epiteliales de la nariz y llegar a los receptores que enviarán la señal al cerebro, indicando la sensación del olor. Un punto importante que deben cumplir las moléculas olorosas es tener un peso molecular bajo, aunque se ha generado cierta controversia en este punto puesto que hay moléculas, tales como los esteroides, que poseen olor aunsiendo de gran tamaño. Otro aspecto en relación con el tamaño de la molécula es su influencia sobre el mecanismo de la quimiorrecepción, pues parece ser que cier tas incapacidades para percibir aromas (anosmias) en los humanos aumentan directamente conforme al tamaño molecular, lo que puede deberse a que las moléculas muy grandes no tienen un buen acomodo en el sitio receptor.
¿Cómo percibimos los olores?
La percepción del olfato involucra tres aspectos importantes: la intensidad, la descripción cualitativa y la apreciación del aroma. La relación entre la concentración de un olor y la intensidad percibida es bien conocida y sigue una distribución logarítmica común, al igual que la de otros sistemas sensoriales. El umbral de detección del olor, definido como la concentración mínima en que se percibe el estímulo, puede ser hasta de partes por trillón para algunos aromas, pero esta estimación varía de un aroma a otro hasta en unas cincuenta veces o más.
Cuando la descripción cualitativa se compara con la intensidad, se presenta una ligera complicación; se cree que los humanos tienen la capacidad de diferenciar hasta 10 mil olores diferentes, pero la mayoría de nosotros solamente percibimos una fracción de ellos a lo largo de la vida, por lo que a menudo la descripción de un olor varía de persona a persona.
En la actualidad se conocen miles de olores; sin embargo, la apreciación de un aroma es subjetiva, ya que se ve influenciada por aspectos culturales o emocionales particulares, es decir, de cada sujeto; de hecho, para algunas personas el mismo perfume puede ser agradable, desagradable o indiferente. No obstante, los estudios fisiológicos enfocados a esta área, basados en pruebas psicológicas, sostienen que muchas sensaciones olfatorias dependen de unas cuantas emociones primarias. Por ello –independientemente de la forma particular en la que cada individuo percibe los olores–, se ha propuesto una sencilla clasificación que postula siete aromas primarios: etéreo, alcanforado, almizcle, floral, mentolado, picante y pútrido. Aunque ésta ha sido la tipificación usualmente aceptada por los científicos, hasta ahora no hay un “mapa de olores” que se ajuste a todos los individuos y que se utilice por las industrias de la cosmética, perfumería y alimentaria. Además, algunos reportes indican que las sensaciones primarias pueden ser hasta cincuenta respuestas o más; sin embargo, es probable que a pesar de que esa cantidad sea significativa, las que puede experimentar el ser humano son solamente las propuestas arriba, argumentando que se posee una cierta “ceguera” para el olor o una ausencia de receptores adecuados para la identificación y caracterización de la molécula que causa tal sensación.
El sentido del olfato
El sentido del olfato es una de las ventanas más sofisticadas a través de la cual un individuo capta información del ambiente externo y por la cual los seres vivos de cualquier especie son capaces de modificar su comportamiento y establecer su gusto por determinados ambientes y alimentos.
En el caso de los humanos, el aparato olfativo tiene varias diferencias respecto del que poseen animales e insectos; incluso se puede decir que nuestro sentido del olfato es extremadamente rudimentario comparado con el de otras especies.
Probablemente esta curiosa diferencia se deba a que la especie humana ha alcanzado a lo largo de su evolución un desarrollo más acentuado de los demás sentidos y también de otras aptitudes, relegando en cierta forma la capacidad olfativa, dando razón de por qué nosotros tenemos otras forma de comunicación distintas de un “simple” sendero aromático.
Paradójicamente, el sentido del olfato de todos los mamíferos conserva ciertas similitudes en la cavidad nasal, tales como su localización y asociación con las vías respiratorias superiores, puesto que los compuestos olorosos se hallan dispersos en el aire, y es así como logran llegar a la nariz para alcanzar las células olfatorias y estimularlas.
Por su parte, la cavidad nasal se encuentra protegida en su totalidad por una capa de células epiteliales y, curiosamente, sólo alrededor de 5% del epitelio que la cubre está relacionada con la función de detectar e identificar los aromas. El epitelio olfativo mide 2.5 cm2 y contiene aproximadamente cincuenta millones de células, mismas que albergan los receptores químicos responsables de la identificación de los olores; dichas células son estructuras provenientes del sistema nervioso central.
Resulta interesante que el epitelio olfativo esté constituido de tres capas principales de células: las células de soporte, que tienen la función de aislar eléctricamente las neuronas; las células receptoras del olfato, que son las responsables de la transmisión del estímulo, y las células basales, que se encuentran en la parte más recóndita del epitelio. Se puede decir entonces que el sentido del olfato no es más que una serie de experiencias dependientes del funcionamiento cerebral y que opera siempre mediante mensajes neuronales.
Algunos científicos consideran que el olor es una percepción que no puede ser medida, en la que el sistema límbico (emocional) es el que afecta la apreciación para clasificar el agrado o desagrado por los aromas, además de que es inf luido por algunos aspectos de orden f isiológico. Tomemos como ejemplo la comida. Cuando se está cocinando o con hambre, el olor tiene una percepción grata; sin embargo, se vuelve desagradable cuando ya se ha comido.
El mecanismo bioquímico del sentido del olfato
A diferencia de la vista, que se explica en función de las leyes fundamentales de la óptica, o de la audición, entendida por fenómenos armónicos, el sentido del olfato tiene aún ciertas incógnitas. La percepción de los aromas, así como el sistema olfativo en general, es de los mecanismos biológicos menos estudiados, probablemente por su localización y, por ende, por su dificultad para reproducirse en un laboratorio. Pese a ello, en el campo de la bioquímica se especula que la generación del impulso nervioso como respuesta a una molécula capaz de producir un aroma es el resultado de una variación de voltaje en la membrana celular, originado por una alteración en los niveles de monofosfato de adenosina cíclico o inositol trifosfato, proceso químico y eléctrico bien conocido que permite postular tres mecanismos hipotéticos mediante los cuales se puede llevar a cabo la traducción del mensaje olfativo: a) por la apertura directa de los canales iónicos alcontacto con la molécula del olor, b) por la alteración de la fluidez de la membrana al interactuar los receptores con los aromas, y c) por una transmisión indirecta del olor, a través de proteínas receptoras ligadas a rutas, dependiente de segundos mensajeros.
Algunos estudios recientes han revelado que el código de olores es diferenciado por múltiples receptores, y que diferentes olores se reconocen por sus distintas combinaciones, de modo que la selección no es específica, sino que se sitúa en un nivel molecular; a tal selección se le ha denominado reconocimiento odotópico. Tal hipótesis ha sido ampliamente aceptada por dos sencillas razones: la primera es que no podrían existir millones de proteínas receptoras, una para cada olor, y la segunda se basa en estudios electrofisiológicos en los que se ha encontrado que una neurona olfativa responde a otras moléculas olorosas de estructura química diferente.
Adicionalmente se ha sugerido que existe agonismo o antagonismo entre los olores y sus receptores en el epitelio olfativo; así, hay una competencia entre las combinaciones de aromas frente a los agonistas para activar un receptor neuronal, o entre agonistas y antagonistas para bloquear una neurona. La incapacidad de un agonista para fijarse en un sitio receptor y activar lo consti tuye uno de los mecanismos más efectivos para provocar una disminución en la percepción del olor; dicha reducción puede considerarse como supresión del olor, y es lo que comúnmente se pretende hacer mediante cier tas formulaciones que
enmascaran los sabores y olores desagradables, como ciertos alimentos, medicinas, cosméticos o productos de limpieza.
Teorías del olfato
Se han propuesto diversas teorías para comprender el funcionamiento biológico de diferenciación del sentido del olfato, tratando de dar una explicación coherente y plausible del modo en que interactúan los olores y los receptores olfativos; sin embargo, debido a la carencia de conocimiento en este campo, muchas de ellas han fracasado o no han sido adecuadamente demostradas. La teoría más aceptada para tal fin es la teoría estereoquímica, que involucra la relación de la estructura tridimensional de la molécula con las propiedades olfativas de los compuestos. Esta teoría se basa en la idea de que los grupos funcionales presentes en una molécula odorífica tienen distintos tamaños, y que mediante un mecanismo similar al modelo enzimático “llave-cerradura”, se puede reconocer la distribución electrónica de un compuesto asociado a un olor característico, siempre y cuando ambas partes coincidan con el arreglo conformacional y una distribución espacial específica.
Olor y estructura molecular
En el ámbito de la investigación farmacéutica, cosmética y de al imentos, la int roducción de nuevos compuestos con propiedades biológicas activas de interés implica un alto costo, ya que requiere técnicas y métodos de laboratorio que, además, necesitan procesos de control y regulación. Por ello se ha venido desarrollando recientemente una metodología para el estudio de nuevos compuestos con cualidades incluso personalizadas, pero que evita una gran inversión económica, ya que el objetivo se logra a través de métodos computacionales que permiten correlacionar la estructura química con la actividad biológica. Estas técnicas hacen posible un análisis adecuado de la estructura molecular, bajo el precepto de que tal relación implica la pertenencia al mismo tipo de estructura y, por consiguiente, a la misma clase de aroma. Así, el diseño computacional de fragancias reúne en el campo de la investigación las bases de la ciencia, la experiencia y el instinto, que se conjugan como si se tratara de una fantasía creacional. A la fecha hay varias reglas para crear agentes odoríficos basados en la estructura química de las sustancias. A continuación se presentan unos cuantos hallazgos de cómo la estructura puede emplearse para predecir o simplemente especular acerca del aroma que un cierto compuesto puede provocar.
Ambergris
Es una fragancia de origen animal secretada en el estómago o tracto intestinal de la ballena y que se libera al mar en forma de una masa rocosa de color grisáceo a negro. Cuando se expone a la luz, al aire y al agua marina, el material se degrada a un color amarillo cremoso, al mismo tiempo que se oxida su principal componente, el alcohol triterpénico ambreina. De acuerdo con el análisis estructural de las propiedades olfativas se ha logrado establecer que debe haber un anillo de trans-decalina dentro de la molécula para dar esta clase de aroma.
Almendras amargas
Este grupo de olores ha sido el modelo por excelencia en los estudios de la relación estructura-actividad, puesto que tiene un olor bien definido. Las moléculas clásicas poseedoras de dicho aroma son el ácido cianhídrico y el benzaldehído, ambos producidos por la hidrólisis de la amigdalina, que es uno de los componentes de las almendras amargas. Sin embargo, si se compara la estructura de ambos compuestos se puede deducir una gran variabilidad estructural, por lo que después de ciertos estudios computacionales se ha logrado establecer que este aroma debe tener un núcleo aromático; cuando los dobles enlaces se encuentran conjugados con grupos funcionales, el aroma se intensifica.
Alcanforado
Los aromas alcanforados se han popularizado por su presencia en productos tales como los ungüentos para tratar resfriados y en antiguas formulaciones de repelentes de polillas. La molécula líder responsable de este olor es el alcanfor, que se puede obtener de forma natural a través del aceite de madera o en el laboratorio, a partir del pineno.
Floral
Las fragancias florales son de las más interesantes dada su gran aceptación por parte de la mayoría de las personas, sobre todo de las mujeres, quienes dominan la industria del perfume. Se pueden usar cientos de diferentes notas florales, siendo las más importantes la rosa, el jazmín y el lirio de los valles. De estos tres solamente se han realizando análisis de estructuraactividad con los dos últimos, puesto que el perfume de rosas es un olor complejo. No obstante, se ha logrado revelar ciertas características moleculares para los olores florales; por ejemplo, sustituyendo anillos de tamaño mediano con grupos funcionales del tipo del alcohol, aldehído, éster o éter.
Frutal
De las fragancias frutales más utilizadas los ésteres forman parte del grupo de compuestos que más se ha estudiado. Se sabe que la intensidad del olor depende considerablemente de la naturaleza de la porción ácida y alcohólica; para los ésteres alifáticos se ha propuesto que aquellos con más de ocho átomos de carbono tienen aromas frutales intensos, y adicionalmente se ha sugerido que las insaturaciones en la proximidad de la función éster reducen el peculiar tono frutal y agregan sensaciones herbales. Por su parte, los olores a uva, frutas rojas, melón y otros de percepción similar se producen cuando ciertos compuestos azufrados tienen bajas concentraciones; de hecho, muchos compuestos orgánicos volátiles de este tipo se utilizan enormemente en la industria de bebidas y alimentos, y no simplemente para productos derivados de frutas, sino para la carne, pan, papas, cerveza y café.
Herbal
Esta vasta familia de fragancias ha sido objeto de muy pocos estudios que relacionen la estructura con el aroma, probablemente porque este aroma es muy difícil de definir. La nota herbal alude tanto al follaje verde de las hojas, tallos y frutos verdes, como al pasto recién cortado. La principal sustancia vinculada con este olor es el cis-3-hexen-1-ol, muchos de cuyos derivados se han introducido en la industria de los sabores y fragancias desde 1960.
Musk
Los aromas del tipo musk son de los más interesantes en el ámbito de la perfumería por dos razones: su gran capacidad como
fijador y sus propiedades características. Son olores considerados cálidos, sensuales y a la vez naturales. Este aroma está químicamente bien definido y abarca cuatro diferentes grupos: macrocíclos nitrogenados, no nitrogenados, bencenoides y esteroidales.
Maderas
En este grupo se incluyen las fragancias derivadas de los aceites esenciales de varios tipos de árboles, como el cedro y el sándalo, y algunos provenientes de hojas como el pachuli y el vetiver. Generalmente son compuestos bicíclicos o tricíclicos de 12 a 17 carbonos y con un grupo funcional alcohol o éster.
El desarrollo e innovación de las fragancias puede en algunos casos considerarse como un capricho meramente placentero, sin embargo, el estudio racional de la relación estructuraactividad de los olores ha permitido comprender algunas semejanzas estructurales en las moléculas de los aromas.
Quizás el enigma del olor entraña, más allá de la fisiología cognitiva, ciertos procesos de comunicación, comportamientos antropológicos y, sobre todo, un mundo de comunicación química que aún está por descubrirse.
OLFATO
http://www.youtube.com/watch?v=EDmKMJcPj8I
El olfato
En el hombre, el sentido del olfato está menos desarrollado que en muchos animales, quizás porque al contrario que éstos, no depende de él para buscar alimento, hallar pareja o protegerse del enemigo.
Los vapores emitidos por las sustancias olorosas penetran por la parte superior de las cavidades o fosas nasales y, después de disolverse en la humedad de la pituitaria amarilla, actúan químicamente sobre los receptores olfativos. Los impulsos nerviosos que resultan de la activación de los receptores son trasmitidos al bulbo olfatorio y de ahí a la corteza cerebral para la formación de la sensación.
Mediante el acto de olfatear, la dirección de la corriente de aire es dirigida hacia la región olfatoria superior de la cavidad, facilitando la llegada de un mayor número de partículas olorosas hasta los receptores olfativos.
Las sensaciones olfatorias suelen confundirse con las del gusto, ya que ambas son producidas por el mismo estímulo químico. En verdad, varios alimentos son apreciados más por el olor que por el sabor.
El olfato contribuye a la iniciación de los procesos de la digestión. Así, cuando los distintos olores alcanzan el centro olfatorio del cerebro, éste envía al estómago los estímulos adecuados para que comience la producción de jugos digestivos; en este proceso interviene también la visión, de tal forma que ante la presencia de la comida empieza a producirse saliva en la boca, lo que facilita la digestión de los carbohidratos.
De todos los órganos de los sentidos, el olfato se
distingue por la rapidez con que se adapta al estímulo. Ello se debe a que,
cuando las células olfatorias se “han acostumbrado” a un determinado olor,
cesan de transmitirlo al cerebro. Esta facilidad para dejar de percibir un olor
no constituye, sin embargo, una limitación muy seria para la vida del hombre,
puesto que sus adaptaciones no dependen tanto del olfato.
Una persona distingue entre dos mil y cuatro mil olores distintos.
Más detalles
El sentido del olfato permite apreciar el olor de los cuerpos.
No todos los cuerpos poseen olor. Los que lo poseen se llaman odoríferos y los que no tienen olor, inodoros.
Para que un cuerpo posea olor es necesario que emita partículas pequeñísimas que se mezclen con el aire. Esas partículas impresionan las terminaciones del nervio olfatorio.
El olfato, como vimos, reside en las fosas nasales que son dos orificios localizados por detrás de la nariz y encima de la boca. Las fosas nasales están separadas por un tabique cartilaginoso: en su parte anterior y óseo en la, porción posterior. Se encuentran por debajo de la cavidad craneana y en su cara externa se advierten tres salientes llamados cornetes superior, medio, e inferior.
Cada fosa nasal se comunica por una abertura con el exterior. A la entrada de ellas se encuentran pelos gruesos y cortos. El interior está recubierto por una membrana llamada pituitaria.
El olfato
En el hombre, el sentido del olfato está menos desarrollado que en muchos animales, quizás porque al contrario que éstos, no depende de él para buscar alimento, hallar pareja o protegerse del enemigo.
El área de la nariz
humana sensible al olor es de unos pocos centímetros cuadrados, mientras que en
el perro, por ejemplo, recubre la membrana glucosa nasal por completo.
Sin embargo, el
olfato humano es el más sensible de todos nuestros sentidos: unas cuantas
moléculas, es decir, una mínima cantidad de materia, bastan para estimular las
células olfativas.
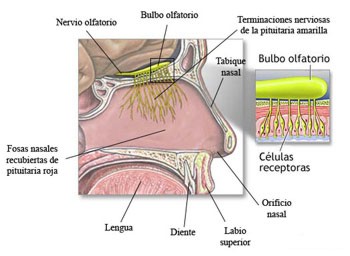
Los receptores olfativos del hombre se encuentran situados en la porción superior de las fosas nasales,
donde la pituitaria amarilla cubre el cornete superior y se comunica con el bulbo olfatorio. (VER
IMAGEN).Los vapores emitidos por las sustancias olorosas penetran por la parte superior de las cavidades o fosas nasales y, después de disolverse en la humedad de la pituitaria amarilla, actúan químicamente sobre los receptores olfativos. Los impulsos nerviosos que resultan de la activación de los receptores son trasmitidos al bulbo olfatorio y de ahí a la corteza cerebral para la formación de la sensación.
Mediante el acto de olfatear, la dirección de la corriente de aire es dirigida hacia la región olfatoria superior de la cavidad, facilitando la llegada de un mayor número de partículas olorosas hasta los receptores olfativos.
Las sensaciones olfatorias suelen confundirse con las del gusto, ya que ambas son producidas por el mismo estímulo químico. En verdad, varios alimentos son apreciados más por el olor que por el sabor.
El olfato contribuye a la iniciación de los procesos de la digestión. Así, cuando los distintos olores alcanzan el centro olfatorio del cerebro, éste envía al estómago los estímulos adecuados para que comience la producción de jugos digestivos; en este proceso interviene también la visión, de tal forma que ante la presencia de la comida empieza a producirse saliva en la boca, lo que facilita la digestión de los carbohidratos.
 |
Una persona distingue entre dos mil y cuatro mil olores distintos.
Más detalles
El sentido del olfato permite apreciar el olor de los cuerpos.
No todos los cuerpos poseen olor. Los que lo poseen se llaman odoríferos y los que no tienen olor, inodoros.
Para que un cuerpo posea olor es necesario que emita partículas pequeñísimas que se mezclen con el aire. Esas partículas impresionan las terminaciones del nervio olfatorio.
El olfato, como vimos, reside en las fosas nasales que son dos orificios localizados por detrás de la nariz y encima de la boca. Las fosas nasales están separadas por un tabique cartilaginoso: en su parte anterior y óseo en la, porción posterior. Se encuentran por debajo de la cavidad craneana y en su cara externa se advierten tres salientes llamados cornetes superior, medio, e inferior.
Cada fosa nasal se comunica por una abertura con el exterior. A la entrada de ellas se encuentran pelos gruesos y cortos. El interior está recubierto por una membrana llamada pituitaria.
Transducción dulce
http://www.youtube.com/watch?v=1Edcmrt2xzc
Las sustancias amargas y dulces se unen a receptores específicos que están acoplados a proteínas G. Las sustancias dulces activan la adenil ciclasa y aumantan la concentración célular de AMP ciclíco. Acto seguido, el AMP ciclíco cierra en canal de potasio, lo que da lugar a una despolarización de la célula gustativa y a la excitación de las eferentes apropiadas.
Las sustancias amargas y dulces se unen a receptores específicos que están acoplados a proteínas G. Las sustancias dulces activan la adenil ciclasa y aumantan la concentración célular de AMP ciclíco. Acto seguido, el AMP ciclíco cierra en canal de potasio, lo que da lugar a una despolarización de la célula gustativa y a la excitación de las eferentes apropiadas.
Transducción amarga
http://www.youtube.com/watch?v=P1OPvHJncJs
Las sustancias amargas activan la fosfolipasa C, y el aumento consiguiente del calcio intracelular da lugar a la liberación de un nuerotransmisor en las eferentes gustativas.
Las sustancias amargas activan la fosfolipasa C, y el aumento consiguiente del calcio intracelular da lugar a la liberación de un nuerotransmisor en las eferentes gustativas.
Transducción salada
http://www.youtube.com/watch?v=P1OPvHJncJs
Las soluciones que son saladas o ácidas activan las células gustativas abriendo un canal iónico específico, que se caracteriza por una alta permeabilidad a los iones de sodio. Este canal es inhibido por una sustancia llamada amilorida. La apertura de este canal iónico despolariza la célula gustativa, lo que da lugar a la excitación de las fibras gustativas eferentes con las que esán conectada. Las soluciones ácidas siempre son de bajo pH y el aumento de la concentración de iones de nitrógeno da lugar al cierre de un canal de sodio. Una vez más, la activación del receptor gustativo da lugar a la despolirización de la célula gustativa. La despolarización abre los canales de calcio dependientes de voltaje, lo que desencadena la exocitosis del neurotransmisor, por parte de las células gustativas, lo que excita a las células nerviosas aferentes apropiadas.
Las soluciones que son saladas o ácidas activan las células gustativas abriendo un canal iónico específico, que se caracteriza por una alta permeabilidad a los iones de sodio. Este canal es inhibido por una sustancia llamada amilorida. La apertura de este canal iónico despolariza la célula gustativa, lo que da lugar a la excitación de las fibras gustativas eferentes con las que esán conectada. Las soluciones ácidas siempre son de bajo pH y el aumento de la concentración de iones de nitrógeno da lugar al cierre de un canal de sodio. Una vez más, la activación del receptor gustativo da lugar a la despolirización de la célula gustativa. La despolarización abre los canales de calcio dependientes de voltaje, lo que desencadena la exocitosis del neurotransmisor, por parte de las células gustativas, lo que excita a las células nerviosas aferentes apropiadas.
Transducción ácida
la transducción ácida se inicia con el aumento de H+, lo cual proboca un
aumento de la conductancia de Na+ y disminución de la concentración de
K+, paso siguido de esto, se lleva a cabo la despolarización de la
célula gustativa, se propicia la secreción de el neurotransmisor por la
célula gustativa, finalmente el resultado es la excitación de la fibra
nerviosa eferente gustativa.
http://www.youtube.com/watch?v=CPNV0dkAQ64
http://www.youtube.com/watch?v=CPNV0dkAQ64
La lengua y los receptores gustativos
http://www.youtube.com/watch?v=JS34YfinA54
Receptores del gusto
Hay cuatro tipos de receptores del gusto: (1) dulce, como los producidos por el azúcar de mesa, (2) agrio, como los producidos por el vinagre, (3) salado, que se producen por la sal de mesa, (4) amargo, producidos por la cafeína o la quinina. Cada uno de estos receptores de sabor está más concentrado en ciertas regiones de la superficie de la lengua. Los receptores dulces están en su mayoría en la punta de la lengua (esto se nota en la preferencia de un niño a lamer una paleta en vez de masticarla). Los receptores agrios se encuentran principalmente a los lados de la lengua y son estimuladas principalmente por los ácidos. Los receptores salados son más comunes en la punta y la parte frontal superior de la lengua, estos son estimulados principalmente por sales inorgánicas. Los receptores inorgánicos se encuentran en la parte posterior de la lengua y son estimuladas por una variedad de sustancias químicas, la mayoría de los cuales son compuestos orgánicos, aunque algunas sales inorgánicas de magnesio y calcio, producen sensaciones muy amargas.GUSTO
http://www.youtube.com/watch?v=7At3zxSnF30
EL GUSTO es el sentido que nos permite saborear las cosas. Se experimenta la sensación del gusto por medio de la lengua.
VIII.1. LA LENGUA HUMANA
La lengua es un órgano muscular, movible, que además de experimentar la sensación del gusto sirve para otras funciones como el habla, el masticamiento y el tragar de los alimentos.
La lengua contiene un conjunto de células especializadas, llamadas yemas gustativas, que son, los órganos especiales del gusto. Además de éstas, la lengua también tiene otro tipo de células que producen saliva, que es necesaria para tragar los alimentos.
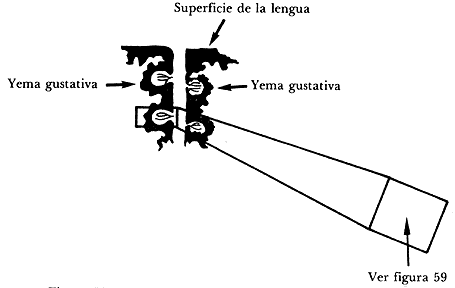
Las yemas gustativas se encuentran hundidas debajo de la
superficie de la lengua en surcos y cavidades (Figura 58). Junto a las
yemas gustativas se encuentran células que secretan líquidos que sirven
para enjuagar las cavidades y los surcos que los ponen en condiciones de
recibir estímulos nuevamente.
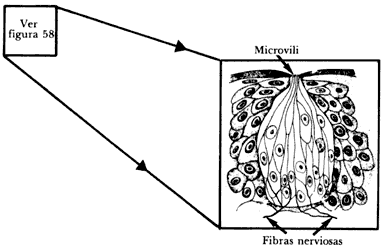
Las yemas gustativas tienen la forma mostrada en la figura 59. Contienen en su interior células en forma de pelos cuyas terminales, los microvili, sobresalen al espacio dentro del surco o cavidad. Las células gustatorias terminan en una fibra nerviosa que comunica las sensaciones recibidas al cerebro.
Las yemas gustativas se encuentran distribuidas de manera no uniforme sobre la superficie de la lengua así como en las estructuras adyacentes de la garganta.
VIII.2. ¿CÓMO PERCIBIMOS LOS SABORES?
En primer lugar, para que la lengua sea sensible a algún objeto es necesario que éste se encuentre húmedo. Si no lo está, entonces se humedecerá con ayuda de la saliva que secreta la misma lengua. Esta es otra función que tiene la saliva.
El objeto ya húmedo se deposita en la lengua y sus moléculas entran en contacto con los microvili, es decir, los extremos de las fibras de las yemas gustativas. De manera análoga a lo que ocurre en el olfato, se lleva a cabo una reacción química que provoca que se desencadene una respuesta de la célula que nos da la sensación del gusto del objeto en cuestión.
Hay que mencionar, además del efecto químico producido en las yemas gustativas, que la sensación del gusto queda determinada por otras propiedades del objeto que producen sensaciones táctiles. Estas propiedades son tanto físicas como químicas. Las primeras de dichas propiedades son el tamaño de la partícula, su textura, su consistencia y su temperatura. Las segundas, las propiedades químicas, producen las sensaciones de frialdad de la menta, lo picoso del chile, etcétera.
La lengua humana no es sensible de manera uniforme a lo largo de su superficie. Así, la parte media no es sensible al gusto como lo son otras partes. En la figura 60 vemos que la punta de la lengua es sensible principalmente a lo dulce. A los lados y en la parte anterior de la lengua se encuentra la sensibilidad a lo salado. La lengua es principalmente sensible a los sabores agrios a los lados, pero en la parte posterior. Finalmente, la parte posterior central de la lengua es sensible principalmente a los sabores amargos.
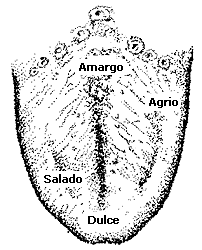
Durante mucho tiempo se creyó que existían cuatro tipos de yemas
gustativas, unas sensibles a lo dulce, otras a lo salado, otras a lo
agrio y otras más a lo amargo. Sin embargo, estudios más detallados han
mostrado que una yema gustativa de la región "dulce", por ejemplo,
también responde a los otros sabores, aunque en menor escala.
La pregunta que uno se puede hacer es ¿cuáles son los factores o propiedades químicas de las sustancias que hacen que las sintamos dulces o amargas? Hasta el momento no se ha encontrado una respuesta definitiva a esta cuestión. No hay una relación directa entre la composición química de la sustancia y el sabor que nos deja al probarla. Sin embargo, sí se han podido determinar algunas propiedades generales que describiremos a continuación
Agrio
Los sabores agrios los percibimos principalmente en sustancias que son ácidas. Estos compuestos contienen átomos de hidrógeno, que son los principales responsables de dicho sabor. Cuando se mezcla la sustancia que contiene el ácido con agua (recuérdese que para que las yemas gustativas reaccionen, las sustancias tienen que estar húmedas), en general desprenden algunos de sus átomos de hidrógeno. Estos átomos de hidrógeno han perdido sus electrones y por lo tanto quedan eléctricamente cargados (Figura 61); se dice que están ionizados. Cuando los átomos de hidrógeno ionizados entran en contacto con las yemas gustativas dan la sensación de un sabor agrio.
Sin embargo, este factor no determina exclusivamente el carácter agrio de una sustancia. Por ejemplo, el ácido cítrico que existe en la naranja, el limón, etc., tiene un sabor agrio muy pronunciado que no se puede explicar con base en la cantidad de iones de hidrógeno que produce. Otro ejemplo es el caso del vinagre, que está compuesto de ácido acético y que es mucho más agrio de lo que se esperaría con base en la cantidad de iones de hidrógeno que produce.
Amargo
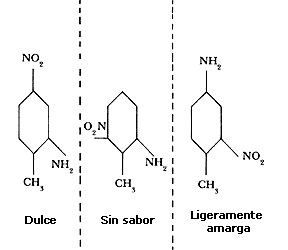
En un buen número de casos, las sustancias amargas son sales inorgánicas de alto peso molecular; esto quiere decir que son moléculas que tienen muchos átomos. Otros casos son moléculas orgánicas que tienen añadidas cadenas carbónicas. Por ejemplo, en la figura 62 se muestran tres moléculas que tienen los mismos átomos pero que difieren en su distribución en el espacio; sin embargo, una de ellas no tiene sabor, la segunda es ligeramente amarga y la tercera es dulce.
Salado
La sal común o de mesa es un compuesto formado por átomos de sodio y de cloro, el cloruro de sodio. Resulta que el sabor salado que da esta sustancia no se debe exclusivamente ni al sodio ni al cloro. Existen sustancias que contienen cloro y no sodio que son saladas, así como compuestos de sodio que no contienen cloro y que también son salados.
Los compuestos llamados sales de bajo peso molecular, es decir, con pocos átomos, tienen sabor predominantemente salado y como se vio arriba, las sales con alto peso molecular son principalmente amargas.
Existen también otras sales que contienen metales. Por ejemplo, sales de mercurio que tienen un sabor metálico mientras que algunas sales de plomo llegan a tener sabor ¡dulce!
Dulce
En general, las sustancias que nos dan la impresión de tener sabor dulce están formadas de compuestos orgánicos como los alcoholes, azúcares, glicoles, etc. La sensación de dulce también se debe a la forma en que estén arreglados los átomos en las moléculas; en la figura 62 vimos compuestos con diferentes disposiciones de los mismos átomos, pero que tienen diferentes sabores, uno de ellos dulce.
En lo que respecta a los sentidos químicos, el olor y el sabor, no se ha avanzado en la misma medida para su comprensión como para los otros sentidos descritos en capítulos anteriores.
VIII.3. ALGUNOS FACTORES QUE AFECTAN LA SENSIBILIDAD DEL GUSTO
A continuación reseñaremos algunos factores que pueden afectar la sensibilidad del gusto.
Uno de estos factores, es la temperatura. Si probamos una sustancia que esté a muy baja temperatura casi no sentiremos su sabor. En general, la lengua será capaz de distinguir sabores si la temperatura de la sustancia es igual o muy parecida a la temperatura del cuerpo.
Otro efecto que nos es muy conocido es el enmascaramiento de sabores. Así, al mezclar azúcar al café se trata de enmascarar el sabor amargo del café. En este caso, la lengua no es capaz de distinguir los sabores individuales de las sustancias mezcladas, es decir, no puede distinguir una mezcla de estímulos gustativos. Esto se contrasta con la capacidad del oído de distinguir una mezcla de estímulos auditivos; por ejemplo, los diferentes instrumentos que tocan al mismo tiempo en una orquesta.
Otra característica del gusto es la adaptación. Si una sustancia ha estado en la boca durante algún tiempo, entonces, en muchas ocasiones, la lengua pierde la sensibilidad a otros sabores y si se prueba otra sustancia dejamos de sentir su sabor o nos da la sensación de que tiene un sabor que no debería tener.
Huelga decir que todavía no se han podido explicar, desde el punto de vista molecular, estos efectos del gusto.
VIII.4. RELACIÓN ENTRE EL OLOR Y EL SABOR
Existe una relación muy íntima entre el olor y el sabor. De hecho cuando comemos un alimento percibimos al mismo tiempo sensaciones en la lengua como en la nariz. Algunas, moléculas del alimento se evaporan y llegan hasta las células olfativas: del gusto y del olfato. Muchas personas que han perdido la capacidad de oler, por ejemplo, debido a algún accidente, dejan de percibir el sabor de la comida.
Hay que mencionar que, en menor escala, también influye en el sabor el sentido de la vista.
EL GUSTO es el sentido que nos permite saborear las cosas. Se experimenta la sensación del gusto por medio de la lengua.
VIII.1. LA LENGUA HUMANA
La lengua es un órgano muscular, movible, que además de experimentar la sensación del gusto sirve para otras funciones como el habla, el masticamiento y el tragar de los alimentos.
La lengua contiene un conjunto de células especializadas, llamadas yemas gustativas, que son, los órganos especiales del gusto. Además de éstas, la lengua también tiene otro tipo de células que producen saliva, que es necesaria para tragar los alimentos.
Las yemas gustativas tienen la forma mostrada en la figura 59. Contienen en su interior células en forma de pelos cuyas terminales, los microvili, sobresalen al espacio dentro del surco o cavidad. Las células gustatorias terminan en una fibra nerviosa que comunica las sensaciones recibidas al cerebro.
Las yemas gustativas se encuentran distribuidas de manera no uniforme sobre la superficie de la lengua así como en las estructuras adyacentes de la garganta.
VIII.2. ¿CÓMO PERCIBIMOS LOS SABORES?
En primer lugar, para que la lengua sea sensible a algún objeto es necesario que éste se encuentre húmedo. Si no lo está, entonces se humedecerá con ayuda de la saliva que secreta la misma lengua. Esta es otra función que tiene la saliva.
El objeto ya húmedo se deposita en la lengua y sus moléculas entran en contacto con los microvili, es decir, los extremos de las fibras de las yemas gustativas. De manera análoga a lo que ocurre en el olfato, se lleva a cabo una reacción química que provoca que se desencadene una respuesta de la célula que nos da la sensación del gusto del objeto en cuestión.
Hay que mencionar, además del efecto químico producido en las yemas gustativas, que la sensación del gusto queda determinada por otras propiedades del objeto que producen sensaciones táctiles. Estas propiedades son tanto físicas como químicas. Las primeras de dichas propiedades son el tamaño de la partícula, su textura, su consistencia y su temperatura. Las segundas, las propiedades químicas, producen las sensaciones de frialdad de la menta, lo picoso del chile, etcétera.
La lengua humana no es sensible de manera uniforme a lo largo de su superficie. Así, la parte media no es sensible al gusto como lo son otras partes. En la figura 60 vemos que la punta de la lengua es sensible principalmente a lo dulce. A los lados y en la parte anterior de la lengua se encuentra la sensibilidad a lo salado. La lengua es principalmente sensible a los sabores agrios a los lados, pero en la parte posterior. Finalmente, la parte posterior central de la lengua es sensible principalmente a los sabores amargos.
La pregunta que uno se puede hacer es ¿cuáles son los factores o propiedades químicas de las sustancias que hacen que las sintamos dulces o amargas? Hasta el momento no se ha encontrado una respuesta definitiva a esta cuestión. No hay una relación directa entre la composición química de la sustancia y el sabor que nos deja al probarla. Sin embargo, sí se han podido determinar algunas propiedades generales que describiremos a continuación
Agrio
Los sabores agrios los percibimos principalmente en sustancias que son ácidas. Estos compuestos contienen átomos de hidrógeno, que son los principales responsables de dicho sabor. Cuando se mezcla la sustancia que contiene el ácido con agua (recuérdese que para que las yemas gustativas reaccionen, las sustancias tienen que estar húmedas), en general desprenden algunos de sus átomos de hidrógeno. Estos átomos de hidrógeno han perdido sus electrones y por lo tanto quedan eléctricamente cargados (Figura 61); se dice que están ionizados. Cuando los átomos de hidrógeno ionizados entran en contacto con las yemas gustativas dan la sensación de un sabor agrio.
Sin embargo, este factor no determina exclusivamente el carácter agrio de una sustancia. Por ejemplo, el ácido cítrico que existe en la naranja, el limón, etc., tiene un sabor agrio muy pronunciado que no se puede explicar con base en la cantidad de iones de hidrógeno que produce. Otro ejemplo es el caso del vinagre, que está compuesto de ácido acético y que es mucho más agrio de lo que se esperaría con base en la cantidad de iones de hidrógeno que produce.
En un buen número de casos, las sustancias amargas son sales inorgánicas de alto peso molecular; esto quiere decir que son moléculas que tienen muchos átomos. Otros casos son moléculas orgánicas que tienen añadidas cadenas carbónicas. Por ejemplo, en la figura 62 se muestran tres moléculas que tienen los mismos átomos pero que difieren en su distribución en el espacio; sin embargo, una de ellas no tiene sabor, la segunda es ligeramente amarga y la tercera es dulce.
Salado
La sal común o de mesa es un compuesto formado por átomos de sodio y de cloro, el cloruro de sodio. Resulta que el sabor salado que da esta sustancia no se debe exclusivamente ni al sodio ni al cloro. Existen sustancias que contienen cloro y no sodio que son saladas, así como compuestos de sodio que no contienen cloro y que también son salados.
Los compuestos llamados sales de bajo peso molecular, es decir, con pocos átomos, tienen sabor predominantemente salado y como se vio arriba, las sales con alto peso molecular son principalmente amargas.
Existen también otras sales que contienen metales. Por ejemplo, sales de mercurio que tienen un sabor metálico mientras que algunas sales de plomo llegan a tener sabor ¡dulce!
Dulce
En general, las sustancias que nos dan la impresión de tener sabor dulce están formadas de compuestos orgánicos como los alcoholes, azúcares, glicoles, etc. La sensación de dulce también se debe a la forma en que estén arreglados los átomos en las moléculas; en la figura 62 vimos compuestos con diferentes disposiciones de los mismos átomos, pero que tienen diferentes sabores, uno de ellos dulce.
Figura 62. Sustancias compuestas de moléculas que tienen los mismos átomos,
pero dispuestos en formas distintas nos producen sabores diferentes.
Hemos de decir que hoy en día todavía no se ha encontrado con
detalle lo que ocurre cuando una sustancia entra en contacto con las
células gustativas de la lengua. En particular, no se tienen respuestas a
preguntas como ¿qué reacciones químicas se llevan a cabo en las yemas
gustativas que desencadenan una señal hacia el cerebro? ¿Qué diferencias
hay, digamos, entre las diversas yemas gustativas que hacen a unas más
sensibles que a otras a un sabor dulce?
En lo que respecta a los sentidos químicos, el olor y el sabor, no se ha avanzado en la misma medida para su comprensión como para los otros sentidos descritos en capítulos anteriores.
VIII.3. ALGUNOS FACTORES QUE AFECTAN LA SENSIBILIDAD DEL GUSTO
A continuación reseñaremos algunos factores que pueden afectar la sensibilidad del gusto.
Uno de estos factores, es la temperatura. Si probamos una sustancia que esté a muy baja temperatura casi no sentiremos su sabor. En general, la lengua será capaz de distinguir sabores si la temperatura de la sustancia es igual o muy parecida a la temperatura del cuerpo.
Otro efecto que nos es muy conocido es el enmascaramiento de sabores. Así, al mezclar azúcar al café se trata de enmascarar el sabor amargo del café. En este caso, la lengua no es capaz de distinguir los sabores individuales de las sustancias mezcladas, es decir, no puede distinguir una mezcla de estímulos gustativos. Esto se contrasta con la capacidad del oído de distinguir una mezcla de estímulos auditivos; por ejemplo, los diferentes instrumentos que tocan al mismo tiempo en una orquesta.
Otra característica del gusto es la adaptación. Si una sustancia ha estado en la boca durante algún tiempo, entonces, en muchas ocasiones, la lengua pierde la sensibilidad a otros sabores y si se prueba otra sustancia dejamos de sentir su sabor o nos da la sensación de que tiene un sabor que no debería tener.
Huelga decir que todavía no se han podido explicar, desde el punto de vista molecular, estos efectos del gusto.
VIII.4. RELACIÓN ENTRE EL OLOR Y EL SABOR
Existe una relación muy íntima entre el olor y el sabor. De hecho cuando comemos un alimento percibimos al mismo tiempo sensaciones en la lengua como en la nariz. Algunas, moléculas del alimento se evaporan y llegan hasta las células olfativas: del gusto y del olfato. Muchas personas que han perdido la capacidad de oler, por ejemplo, debido a algún accidente, dejan de percibir el sabor de la comida.
Hay que mencionar que, en menor escala, también influye en el sabor el sentido de la vista.
Percepción
http://www.youtube.com/watch?v=ym1zscafj8U
La percepción (del latín perceptĭo) consiste en recibir, a través de los sentidos, las imágenes, sonidos, impresiones o sensaciones externas. Se trata de una función psíquica que permite al organismo captar, elaborar e interpretar la información que llega desde el entorno.

Es importante diferenciar entre el estímulo, que pertenece al mundo exterior y genera el primer efecto en la cadena del conocimiento, y la percepción, que es un proceso psicológico y pertenece al mundo interior. Podría decirse que el estímulo es la energía física, mecánica, térmica, química o electromagnética que excita o activa a un receptor sensorial.
La percepción visual es aquella sensación interior de conocimiento aparente, resultante de un estímulo o impresión luminosa registrada por los ojos.
Por lo general, este acto óptico-físico funciona de modo similar en
todas las personas, ya que las diferencias fisiológicas de los órganos
visuales apenas afectan al resultado de la percepción.
Las principales diferencias surgen con la interpretación de la información recibida, a causa de las desigualdades de cultura, educación, inteligencia y edad, por ejemplo. En este sentido, las imágenes pueden “leerse” o interpretarse tal como un texto literario, por lo que existe en la operación de percepción visual la posibilidad de un aprendizaje para profundizar el sentido de la lectura.
Los psicólogos de la Gestalt, a comienzos del siglo XX, fueron los primeros en proponer una teoría filosófica de la forma. Max Wertheimer, Wolfgang Köhler, Kurt Koffka y Kurt Lewin, entre otros, aseguraron que, en la percepción, el todo es mayor que la suma de las partes.
Es importante diferenciar entre el estímulo, que pertenece al mundo exterior y genera el primer efecto en la cadena del conocimiento, y la percepción, que es un proceso psicológico y pertenece al mundo interior. Podría decirse que el estímulo es la energía física, mecánica, térmica, química o electromagnética que excita o activa a un receptor sensorial.
Las principales diferencias surgen con la interpretación de la información recibida, a causa de las desigualdades de cultura, educación, inteligencia y edad, por ejemplo. En este sentido, las imágenes pueden “leerse” o interpretarse tal como un texto literario, por lo que existe en la operación de percepción visual la posibilidad de un aprendizaje para profundizar el sentido de la lectura.
Los psicólogos de la Gestalt, a comienzos del siglo XX, fueron los primeros en proponer una teoría filosófica de la forma. Max Wertheimer, Wolfgang Köhler, Kurt Koffka y Kurt Lewin, entre otros, aseguraron que, en la percepción, el todo es mayor que la suma de las partes.
Visión a color
http://www.youtube.com/watch?v=xK2-WTdQ2Uc
Si una pelota playera roja y brillante viene girando hacia uno, se ve su color, su forma y su movimiento, todo a la vez—pero el cerebro se ocupa de cada una de estas características por separado", explica.
Las neuronas son máquinas de cómputos relativamente lentas, dice Nathans, un investigador del HHMI, en la Facultad de Medicina, en la Universidad Johns Hopkins. "Les lleva varios milisegundos recibir un estímulo y generar otro. Sin embargo, se pueden ver cosas en una fracción de segundo, el tiempo para no más de 100 pasos sucesivos. Por esto mismo, el sistema requiere de un procesamiento paralelo".
El día que se enteró de los nuevos descubrimientos sobre cómo vemos en blanco y negro, Nathans se interesó en cómo vemos en colores. Era 1980, y él era estudiante en la Facultad de Medicina de Stanford, recuerda cuando Lubert Stryer y Denis Baylor, ambos en Stanford, describieron sus notables descubrimientos acerca del funcionamiento de las células de tipo bastón.
Las células de tipo bastón—una de las dos clases de células fotorreceptoras de la retina—nos permiten ver a la luz suave de las estrellas durante una noche brumosa. "Baylor mostró que las células de tipo bastón consiguen lo que es fundamental para la sensibilidad a la luz—poder responder a un único fotón o partícula de luz", dice Nathans. "Fue un experimento hermoso". (El trabajo de Baylor fue realizado en colaboración con Trevor Lamb y King-Wai Yau).
Entonces, Stryer explicó cómo la rodopsina, la proteína receptora sensible a la luz que está presente en las membranas de los discos de las células de tipo bastón, anuncia la llegada de este pequeño pulso de luz a la maquinaria de señalización dentro de la célula. Stryer había encontrado que la rodopsina sólo podía hacer esto con la ayuda de una intermediaria, llamada proteína G, que pertenecía a una familia de proteínas que ya era conocida por los bioquímicos, gracias al estudio que ellos habían hecho sobre cómo las células responden a las hormonas y a los factores del crecimiento.
Inmediatamente, Nathans se dio cuenta que esto significaba que la estructura de la rodopsina misma podía ser similar a la de los receptores de las hormonas. Comenzó a jugar en su mente con las posibilidades. "Y corrí, literalmente corrí, a la biblioteca y empecé a leer sobre la visión", dice.
Hasta entonces, Nathans había estado estudiando la genética de las moscas de la fruta. Pero al leer la publicación del biólogo George Wald, de la Universidad de Harvard—una transcripción de la conferencia de Wald, sobre "La base molecular de la excitación visual", durante la entrega del Premio Nobel de 1967—Nathans tomó un rumbo diferente. Resolvió hacer lo que el mismo Wald había deseado hacer 40 años antes: encontrar las proteínas receptoras de la retina que responden al color.
Las células de tipo bastón sólo permiten la visión en la luz débil y no perciben los colores. "Levántese una noche de luna oscura y mire a su alrededor", sugiere David Hubel de la Facultad de Medicina de Harvard, galardonado con el Premio Nobel por su investigación sobre la visión. "A pesar de que pueda ver algunas formas bastante bien, los colores estarán completamente ausentes. Es notable cómo pocas personas se dan cuenta de que no necesitan de la visión de color para poder ver con luz débil".
Pero la retina humana también contiene otro tipo de célula fotorreceptora: los conos, los cuales operan con luz brillante y son los responsables de la gran agudeza visual, además de la visión de los colores.
Los conos y bastones forman un mosaico desigual dentro de la retina, con bastones que generalmente exceden en número a los conos, en una relación de 10 a 1, excepto en el centro de la retina o fóvea. Los conos están sumamente concentrados en la fóvea, un área que Nathans llama "el milímetro cuadrado de tejido más valioso del cuerpo".
A pesar de que la fóvea es esencial para una vista excelente, es menos sensible a la luz que la retina que la rodea. Por ello, si deseamos detectar una estrella débil en la noche, debemos fijar la vista ligeramente hacia el costado de la estrella para proyectar su imagen sobre los bastones, que son más sensibles, ya que la estrella emite una luz que es insuficiente para activar a un cono.
Si una pelota playera roja y brillante viene girando hacia uno, se ve su color, su forma y su movimiento, todo a la vez—pero el cerebro se ocupa de cada una de estas características por separado", explica.
Las neuronas son máquinas de cómputos relativamente lentas, dice Nathans, un investigador del HHMI, en la Facultad de Medicina, en la Universidad Johns Hopkins. "Les lleva varios milisegundos recibir un estímulo y generar otro. Sin embargo, se pueden ver cosas en una fracción de segundo, el tiempo para no más de 100 pasos sucesivos. Por esto mismo, el sistema requiere de un procesamiento paralelo".
El día que se enteró de los nuevos descubrimientos sobre cómo vemos en blanco y negro, Nathans se interesó en cómo vemos en colores. Era 1980, y él era estudiante en la Facultad de Medicina de Stanford, recuerda cuando Lubert Stryer y Denis Baylor, ambos en Stanford, describieron sus notables descubrimientos acerca del funcionamiento de las células de tipo bastón.
Las células de tipo bastón—una de las dos clases de células fotorreceptoras de la retina—nos permiten ver a la luz suave de las estrellas durante una noche brumosa. "Baylor mostró que las células de tipo bastón consiguen lo que es fundamental para la sensibilidad a la luz—poder responder a un único fotón o partícula de luz", dice Nathans. "Fue un experimento hermoso". (El trabajo de Baylor fue realizado en colaboración con Trevor Lamb y King-Wai Yau).
Entonces, Stryer explicó cómo la rodopsina, la proteína receptora sensible a la luz que está presente en las membranas de los discos de las células de tipo bastón, anuncia la llegada de este pequeño pulso de luz a la maquinaria de señalización dentro de la célula. Stryer había encontrado que la rodopsina sólo podía hacer esto con la ayuda de una intermediaria, llamada proteína G, que pertenecía a una familia de proteínas que ya era conocida por los bioquímicos, gracias al estudio que ellos habían hecho sobre cómo las células responden a las hormonas y a los factores del crecimiento.
Inmediatamente, Nathans se dio cuenta que esto significaba que la estructura de la rodopsina misma podía ser similar a la de los receptores de las hormonas. Comenzó a jugar en su mente con las posibilidades. "Y corrí, literalmente corrí, a la biblioteca y empecé a leer sobre la visión", dice.
Hasta entonces, Nathans había estado estudiando la genética de las moscas de la fruta. Pero al leer la publicación del biólogo George Wald, de la Universidad de Harvard—una transcripción de la conferencia de Wald, sobre "La base molecular de la excitación visual", durante la entrega del Premio Nobel de 1967—Nathans tomó un rumbo diferente. Resolvió hacer lo que el mismo Wald había deseado hacer 40 años antes: encontrar las proteínas receptoras de la retina que responden al color.
Las células de tipo bastón sólo permiten la visión en la luz débil y no perciben los colores. "Levántese una noche de luna oscura y mire a su alrededor", sugiere David Hubel de la Facultad de Medicina de Harvard, galardonado con el Premio Nobel por su investigación sobre la visión. "A pesar de que pueda ver algunas formas bastante bien, los colores estarán completamente ausentes. Es notable cómo pocas personas se dan cuenta de que no necesitan de la visión de color para poder ver con luz débil".
Pero la retina humana también contiene otro tipo de célula fotorreceptora: los conos, los cuales operan con luz brillante y son los responsables de la gran agudeza visual, además de la visión de los colores.
Los conos y bastones forman un mosaico desigual dentro de la retina, con bastones que generalmente exceden en número a los conos, en una relación de 10 a 1, excepto en el centro de la retina o fóvea. Los conos están sumamente concentrados en la fóvea, un área que Nathans llama "el milímetro cuadrado de tejido más valioso del cuerpo".
A pesar de que la fóvea es esencial para una vista excelente, es menos sensible a la luz que la retina que la rodea. Por ello, si deseamos detectar una estrella débil en la noche, debemos fijar la vista ligeramente hacia el costado de la estrella para proyectar su imagen sobre los bastones, que son más sensibles, ya que la estrella emite una luz que es insuficiente para activar a un cono.
sábado, 7 de julio de 2012
fotoquímica de la visión
http://www.youtube.com/watch?v=pgKJzUBxX9s
Cuando la retina esta en condiciones de oscuridad, se encuentran abiertos una serie de canales iónicos a nivel de los segmentos externos de los fotorreceptores que permiten la entrada fundamentalmente de iones Sodio. Esta entrada de Sodio, despolariza parcialmente a los fotorreceptores, permitiendo la liberación de neurotransmisor a nivel de sus terminales sinápticos.
El transmisor liberado se supone que es Glutamato. Cuando la luz estimula a la molécula de rodopsina, se producen una sería de cambios que se presentan esquemáticamente en la imagen siguiente, que van a producir el cierre de los canales iónicos permeables al sodio.
Por tanto cesa la entrada de sodio y el fotorreceptor se hiperpolariza, con lo que deja de liberar el neurotransmisor.
La corriente que se produce durante las condiciones de oscuridad es debida en un 80% a la entrada de iones sodio, sin embargo el canal es también permeable a los iones calcio y magnesio. Además en oscuridad debe existir un mecanismo para eliminar tanto el calcio como el exceso de sodio. Este mecanismo parece ser que consiste en un intercambiador sodio/calcio a nivel de la membrana de los segmentos externo. El calcio, además tiene un importante papel en todo el proceso de la fototransducción, ya que aunque no participa directamente en la cascada de la fototransducción, mejora la capacidad de los bastones para recuperarse después de la iluminación, teniendo un importante papel regulador en los fenómenos de adaptación a las condiciones de luz/oscuridad.
Cuando la retina esta en condiciones de oscuridad, se encuentran abiertos una serie de canales iónicos a nivel de los segmentos externos de los fotorreceptores que permiten la entrada fundamentalmente de iones Sodio. Esta entrada de Sodio, despolariza parcialmente a los fotorreceptores, permitiendo la liberación de neurotransmisor a nivel de sus terminales sinápticos.
El transmisor liberado se supone que es Glutamato. Cuando la luz estimula a la molécula de rodopsina, se producen una sería de cambios que se presentan esquemáticamente en la imagen siguiente, que van a producir el cierre de los canales iónicos permeables al sodio.
Por tanto cesa la entrada de sodio y el fotorreceptor se hiperpolariza, con lo que deja de liberar el neurotransmisor.
La corriente que se produce durante las condiciones de oscuridad es debida en un 80% a la entrada de iones sodio, sin embargo el canal es también permeable a los iones calcio y magnesio. Además en oscuridad debe existir un mecanismo para eliminar tanto el calcio como el exceso de sodio. Este mecanismo parece ser que consiste en un intercambiador sodio/calcio a nivel de la membrana de los segmentos externo. El calcio, además tiene un importante papel en todo el proceso de la fototransducción, ya que aunque no participa directamente en la cascada de la fototransducción, mejora la capacidad de los bastones para recuperarse después de la iluminación, teniendo un importante papel regulador en los fenómenos de adaptación a las condiciones de luz/oscuridad.
El ojo y las células fotoreceptoras
http://www.youtube.com/watch?feature=endscreen&NR=1&v=PZEsR2hq4Qk
TUNICA NERVIOSA La retina, la tercera túnica y la más interna del ojo, es la porción neural que contiene las células fotorreceptoras conocidas como bastones y conos. La retina está formada por una capa pigmentada. La porción neural de la retina se forma a partir de la capa interna de la copa óptica y es la retina propiamente dicha.
4 El disco óptico, situado en la pared posterior del ojo, es el sitio de salida del nervio óptico, es insensible a la luz y; por esta razón, se conoce como punto ciego de la retina Aproximadamente a 2.5 mm a un lado del disco óptico se halla una zona de color amarillo en la pared de la retina llamada mácula lútea (punto amarillo). Localizada en el centro de esta mancha existe una depresión oval, la fóvea central, en donde es mayor la agudeza visual. La fóvea es un área especializada de la retina que sólo contiene conos. La porción de la retina que actúa en la fotorrecepción reviste la superficie interna de la capa coroides, desde el disco óptico hasta la ora serrata y se integra con 10 capas distintas. Desde la parte exterior a la interior, estas capas son las siguientes:
1. Epitelio pigmentario.
2. Capa de conos y bastones.
3. Membrana limitante externa.
4. Capa nuclear externa.
5. Capa plexiforme externa.
6. Capa nuclear interna.
7. Capa plexiforme interna.
8. Capa de células ganglionares.
9. Capa de fibras del nervio óptico.
10. Membrana limitante interna.
Epitelio pigmentario. El epitelio pigmentario que deriva de la capa externa de la copa óptica, está compuesto de células cuboides a cilíndricas. Estas células están unidas a la membrana de Bruch, situada entre la coroides y las células de pigmento. Los vértices de las células muestran microvellosidades y estructuras similares a manguillos que rodean y aíslan las puntas de células fotorreceptoras individuales. La característica más distintiva de las células pigmentarias es su abundancia en gránulos de melanina, que sintetizan y almacenan estas células en sus porciones apicales. El epitelio pigmentario tiene varias funciones. Las células epiteliales pigmentarias absorben la luz una vez que pasa a través de los fotorreceptores. Estas células pigmentarias fagocitan continuamente discos membranosos gastados de las puntas de los bastones fotorreceptores. También tienen una función activa en la visión al esterificar los derivados de la vitamina A.
Capa de conos y bastones. La porción óptica de la retina aloja dos tipos distintos de células fotorrepectoras llamadas conos y bastones, son células polarizadas cuyas porciones apicales, conocidas como segmentos externos, son dendritas especializadas. Las bases de los bastones y conos hacen sinapsis con las células subyacentes de la capa bipolar. Los bastones son receptores especializados para la luz tenue; los conos son receptores especializados para la recepción de luz brillante. Los conos están separados adicionalmente para la visión a colores, mientras que los bastones sólo perciben luz.
* Ya que los conos están altamente
TUNICA NERVIOSA La retina, la tercera túnica y la más interna del ojo, es la porción neural que contiene las células fotorreceptoras conocidas como bastones y conos. La retina está formada por una capa pigmentada. La porción neural de la retina se forma a partir de la capa interna de la copa óptica y es la retina propiamente dicha.
4 El disco óptico, situado en la pared posterior del ojo, es el sitio de salida del nervio óptico, es insensible a la luz y; por esta razón, se conoce como punto ciego de la retina Aproximadamente a 2.5 mm a un lado del disco óptico se halla una zona de color amarillo en la pared de la retina llamada mácula lútea (punto amarillo). Localizada en el centro de esta mancha existe una depresión oval, la fóvea central, en donde es mayor la agudeza visual. La fóvea es un área especializada de la retina que sólo contiene conos. La porción de la retina que actúa en la fotorrecepción reviste la superficie interna de la capa coroides, desde el disco óptico hasta la ora serrata y se integra con 10 capas distintas. Desde la parte exterior a la interior, estas capas son las siguientes:
1. Epitelio pigmentario.
2. Capa de conos y bastones.
3. Membrana limitante externa.
4. Capa nuclear externa.
5. Capa plexiforme externa.
6. Capa nuclear interna.
7. Capa plexiforme interna.
8. Capa de células ganglionares.
9. Capa de fibras del nervio óptico.
10. Membrana limitante interna.
Epitelio pigmentario. El epitelio pigmentario que deriva de la capa externa de la copa óptica, está compuesto de células cuboides a cilíndricas. Estas células están unidas a la membrana de Bruch, situada entre la coroides y las células de pigmento. Los vértices de las células muestran microvellosidades y estructuras similares a manguillos que rodean y aíslan las puntas de células fotorreceptoras individuales. La característica más distintiva de las células pigmentarias es su abundancia en gránulos de melanina, que sintetizan y almacenan estas células en sus porciones apicales. El epitelio pigmentario tiene varias funciones. Las células epiteliales pigmentarias absorben la luz una vez que pasa a través de los fotorreceptores. Estas células pigmentarias fagocitan continuamente discos membranosos gastados de las puntas de los bastones fotorreceptores. También tienen una función activa en la visión al esterificar los derivados de la vitamina A.
Capa de conos y bastones. La porción óptica de la retina aloja dos tipos distintos de células fotorrepectoras llamadas conos y bastones, son células polarizadas cuyas porciones apicales, conocidas como segmentos externos, son dendritas especializadas. Las bases de los bastones y conos hacen sinapsis con las células subyacentes de la capa bipolar. Los bastones son receptores especializados para la luz tenue; los conos son receptores especializados para la recepción de luz brillante. Los conos están separados adicionalmente para la visión a colores, mientras que los bastones sólo perciben luz.
* Ya que los conos están altamente
Luz visible
http://www.youtube.com/watch?v=hbZdmxJj3uE
Las ondas de luz tienen longitudes de onda entre 400 y 700 nanómetros (4 000 y 7 000 Å). A medida que el arcoiris se llena de matices, nuestros ojos perciben diferentes longitudes de ondas de luz. La luz roja tiene longitudes de onda relativamente largas, aproximadamente 700 nm (10-9 metros) de largo. La luz azul y la luz morada tienen ondas cortas, aproximadamente 400 nm. Las ondas más cortas vibran a mayores frecuencias, y tienen energías más elevadas. Las luz roja tiene una frecuencia aproximada de 430 terahertz, mientras que la frecuencia de la luz azul es de aproximadamente 750 terahertz. Los fotones rojos tienen aproximadamente 1.8 electrón-Volt(eV) de energía, mientras que cada fotón azul transmite aproximadamente 3.1 eV.
Los vecinos de la luz visible en el espectro EM son la radiación infrarroja de un lado, y luz ultravioleta del otro lado. La radiación infrarroja tiene longitudes de ondas más largas que la luz roja, es por esto que oscila a una frecuencia menor y lleva consigo menor energía. La radiación ultravioleta tiene longitudes de ondas más cortas que la luz azul o violeta, por lo que oscila más rápidamente, y porta mayor cantidad de energía por protón que la luz visible.
La luz viaja a la increíble velocidad de 299 792 458 kilometros por segundo (aproximadamente 186 282.4 millas por segundo). A esta increíble velocidad, ¡la luz podría girar más de siete veces alrededor de la Tierra en cada segundo!. La letra "c" minúscula se usa en las ecuaciones para representar la velocidad de la luz, como es el caso de la famosa relación entre energía y materia de Einstein: "E = mc2". Todas las formas de ondas electromagnéticas, incluyendo los rayos X y las ondas de radio , y todas las demás frecuencias a lo largo del espectro EM, también viajan a la velocidad de la luz. La luz viaja más rapidamente en el vacío, y se mueve más lentamente en materiales como agua o vídrio.
Luz Visible
La luz visible es una de las formas como se desplaza la energía. Las ondas de luz son el resultado de vibraciones de campos eléctricos y magnéticos, y es por esto que son una forma de radiación electromagnética (EM). La luz visible es tan sólo uno de los muchos tipos de radiación EM, y ocupa un pequeño rango de la totalidad del espectro electromagnético . Sin embargo, podemos percibir la luz directamente con nuestros ojos, y por la gran importancia que tiene para nosotros, elevamos la importancia de esta pequeña ventana en el espectro de rayos EM.Las ondas de luz tienen longitudes de onda entre 400 y 700 nanómetros (4 000 y 7 000 Å). A medida que el arcoiris se llena de matices, nuestros ojos perciben diferentes longitudes de ondas de luz. La luz roja tiene longitudes de onda relativamente largas, aproximadamente 700 nm (10-9 metros) de largo. La luz azul y la luz morada tienen ondas cortas, aproximadamente 400 nm. Las ondas más cortas vibran a mayores frecuencias, y tienen energías más elevadas. Las luz roja tiene una frecuencia aproximada de 430 terahertz, mientras que la frecuencia de la luz azul es de aproximadamente 750 terahertz. Los fotones rojos tienen aproximadamente 1.8 electrón-Volt(eV) de energía, mientras que cada fotón azul transmite aproximadamente 3.1 eV.
Los vecinos de la luz visible en el espectro EM son la radiación infrarroja de un lado, y luz ultravioleta del otro lado. La radiación infrarroja tiene longitudes de ondas más largas que la luz roja, es por esto que oscila a una frecuencia menor y lleva consigo menor energía. La radiación ultravioleta tiene longitudes de ondas más cortas que la luz azul o violeta, por lo que oscila más rápidamente, y porta mayor cantidad de energía por protón que la luz visible.
La luz viaja a la increíble velocidad de 299 792 458 kilometros por segundo (aproximadamente 186 282.4 millas por segundo). A esta increíble velocidad, ¡la luz podría girar más de siete veces alrededor de la Tierra en cada segundo!. La letra "c" minúscula se usa en las ecuaciones para representar la velocidad de la luz, como es el caso de la famosa relación entre energía y materia de Einstein: "E = mc2". Todas las formas de ondas electromagnéticas, incluyendo los rayos X y las ondas de radio , y todas las demás frecuencias a lo largo del espectro EM, también viajan a la velocidad de la luz. La luz viaja más rapidamente en el vacío, y se mueve más lentamente en materiales como agua o vídrio.
VISION
http://www.youtube.com/watch?v=mvBoWKe6QK0
Sentido de la vista o de la visión
El ojo es el órgano de visión. El ojo tiene una estructura compleja que consiste de un lente que enfoca la luz en la retina. La retina esta cubierta por dos tipos de células fotorreceptoras con formas de bastones y conos. Las células en forma de cono son sensitivas al color de la luz y están situadas en la parte de la retina llamada la fóvea, donde el lente enfoca la luz. Las células en forma de bastón no son sensitivas al color, pero tienen elevada sensibilidad a la luz. Los bastones están situados alrededor de la fóvea y son responsables por la visión periférica y la visión nocturna. El ojo está conectado al cerebro a través del nervio óptico. El punto de esta conexión se llama papila o "punto ciego" porque es insensible a la luz. Experimentos científicos han demostrado que la parte posterior del cerebro corresponde con la percepción visual en la retina.El cerebro combina las dos imágenes percibidas por nuestros ojos en una sola imagen tridimensional. Aunque la imagen en la retina esta invertida por la acción del lente, el cerebro la rectifica y percibimos los objetos en su posición original. Algunos experimentos con anteojos que invierten las imágenes por medio de prismas han demostrado que los sujetos pasan por un período inicial de gran confusión, pero eventualmente perciben las imágenes de forma correcta.
La sensibilidad del ojo humano es fenomenal. En la obscuridad, los bastones secretan un fotopigmento llamado rodopsina que aumenta la fotorecepción y hace posible detectar niveles muy bajos de luz. En luz intensa, el iris se contrae reduciendo el tamaño de la abertura que admite luz al ojo y una sustancia obscura reduce la exposición y protege a las células de la retina. El ojo tiene sensibilidad al espectro de la luz desde el rojo hasta el violeta. Las frecuencias electromagnéticas más bajas en el infrarrojo se detectan como calor, pero no se pueden ver. Frecuencias más altas en el ultravioleta y más allá tampoco se pueden ver, pero se pueden detectar como comezón en la piel o en los ojos dependiendo de la frecuencia. El ojo humano no es sensible a la polarización de la luz, es decir, a la luz formada por fotones cuyos vectores de campo eléctrico están alineados en la misma dirección. Mientras tanto, las abejas son sensibles a la luz polarizada, y tienen una gama visual que se extiende hasta la luz ultravioleta. Algunas clases de serpientes tienen sensores infrarrojos que les permiten cazar en oscuridad absoluta usando solamente la radiación emitida por el calor de sus presas. Los pájaros tienen una mayor densidad de células fotoreceptoras que los seres humanos en sus retinas, y por eso poseen mejor agudeza visual.
El daltonismo o discromatopsia es un defecto genético que hace imposible distinguir los colores correctamente. Personas con ciertos tipos de daltonismo confundan el verde y el rojo. Esto puede ser una desventaja para ciertos tipos de ocupaciones. Para una persona daltónica, una persona con visión normal puede parecer tener percepción extrasensorial, pero este término tiene que reservarse para habilidades más allá de la gama normal.
Cuantificación del transporte
http://www.youtube.com/watch?v=mozX-Fvuns8
la cuantificacion del trasporte, se lleva a acno a traves de los productos que producen, de acuerdo al producto que produce, determinado tipo de transporte, y determinando asi su producto y cuantificarlo por distitas tecnicas, sgeun sea el caso
la cuantificacion del trasporte, se lleva a acno a traves de los productos que producen, de acuerdo al producto que produce, determinado tipo de transporte, y determinando asi su producto y cuantificarlo por distitas tecnicas, sgeun sea el caso
Sistemas de transporte
http://www.youtube.com/watch?v=UpOgVFgjdZI
Lo primero que tiene que hacer un microorganismo a la hora de su nutrición
es captar los nutrientes que necesite desde el medio exterior. Debido a que la
bicapapa lipídica actúa como barrera que impide el paso de la mayor parte de
las sustancias, esto significa que deben existir mecanismos específicos para
lograr la entrada de los nutrientes. Además, teniendo en cuenta que las
bacterias suelen vivir en medios diluidos, deben realizar un “trabajo” para
trasladar muchos de esos nutrientes en contra del gradiente de concentración.
Tradicionalmente se viene considerando tres métodos principales de
transporte de sustancias a través de la membrana:
|
transporte pasivo inespecífico (=
difusión simple);
| |
|
transporte pasivo específico (=
difusión facilitada);
| |
|
transporte activo.
|
Como veremos, los más
importantes en procariotas son los sistemas de transporte activo.
2.1 TRANSPORTE PASIVO INESPECÍFICO O DIFUSIÓN SIMPLE
Este transporte consiste en la difusión pasiva de ciertas sustancias
para las que la membrana es impermeable, debido a la diferencia de concentración
(DC)
a ambos lados de dicha membrana (la sustancia tiene mayor concentración fuera
que dentro de la célula). Aparte de esta diferencia de concentración, en la
difusión pasiva influyen:
|
la constante de permeabilidad (P),
es decir, el grado de permeabilidad de la membrana a la sustancia en cuestión;
| |
|
el área o superficie total (A) a
través de la que se produce el transporte.
|
Las membranas citoplásmicas son impermeables en sí mismas a la mayor
parte de las moléculas. Sólo se da en el caso de O2, CO2,
NH3, agua y otras pequeñas sustancias polares no ionizadas.
La difusión simple se produce por el paso de estas sustancias a través
de poros inespecíficos de la membrana citoplásmica.
2.2 TRANSPORTE PASIVO ESPECÍFICO O DIFUSIÓN FACILITADA
Es un proceso que permite el paso de compuestos por difusión a través
de transportadores estereoespecíficos y (al igual que en el caso anterior)
sobre la base de un gradiente de concentración (en la dirección termodinámicamente
favorable).
El transportador suele ser una proteína integral de membrana (permeasa o facilitador), cuya
conformación determina un canal interior, y por el cual un determinado sustrato
puede alcanzar el interior, sin gasto de energía. Se piensa que cuando el
soluto se une a la parte de la permeasa que da al exterior, esta proteína sufre
un cambio conformacional que libera la molécula en el interior. Como al entrar
la molécula, enseguida entra en el metabolismo y desaparece como tal, esto
basta para mantener el gradiente de concentración que permite esta difusión.
La difusión facilitada exhibe propiedades similares a las de las reacciones
enzimáticas:
|
Especificidad de sustrato: cada
permeasa transporte un solo tipo de sustratos químicamente parecidos.
| |
|
Cinética de saturación de tipo
Michaelis-Menten, es decir, la velocidad de transporte aumenta con la
concentración de sustrato, hasta un valor límite (Vmax) por
encima del cual ulteriores aumentos del soluto no aumentan dicha velocidad
(debido a que todas las porinas disponibles están ya totalmente ocupadas):
|
Velocidad
de entrada: Vent = Vmáx
· [Sext] /Km
+ [Sext]
Velocidad
de salida: Vsal
= Vmáx · [Sint]
/Km + [Sint]
Aunque este sistema de transporte es muy común en eucariotas, es muy
raro encontrarlo en bacterias. La explicación evolutiva es que los procariotas
suelen vivir en ambientes con pocas concentraciones de nutrientes, y por lo
tanto no es frecuente que se den gradientes adecuados. Una de las pocas
excepciones la constituye el glicerol, que es transportado por difusión
facilitada en una amplia gama de bacterias, tanto Gram-positivas como
Gram-negativas. Conforme el glicerol entra, es rápidamente convertido a
glicerol-fosfato; por lo tanto, la concentración interna de glicerol como tal
es prácticamente nula, lo que facilita esta difusión incluso a bajas
concentraciones exteriores de esta sustancia. En Zymomonas
existe un facilitador de membrana que transporta glucosa.
2.3 TRANSPORTE ACTIVO
Consiste en el transporte de sustancias en
contra de un gradiente de concentración, lo que requiere un gasto energético. En la mayor parte de
los casos este transporte activo (que supone un trabajo osmótico) se realiza
|
a
expensas de un gradiente de H+ (potencial electroquímico de
protones) previamente creado a ambos
lados de la membrana, por procesos de respiración y fotosíntesis;
| |
|
por
hidrólisis de ATP.
|
Los sistemas de transporte activo son los más abundantes entre las
bacterias, y se han seleccionado evolutivamente debido a que en sus medios
naturales la mayoría de los procariotas se encuentran de forma permanente o
transitoria con una baja concentración de nutrientes.
Los sistemas de transporte activo están basados en permeasas específicas
e inducibles. El modo en que se
acopla la energía metabólica con el transporte del soluto aún no está
dilucidado, pero en general se maneja la hipótesis de que las permeasas, una
vez captado el sustrato con gran afinidad, experimentan un cambio conformacional
dependiente de energía que les hace perder dicha afinidad, lo que supone la
liberación de la sustancia al interior celular.
Estudiaremos los siguientes tipos de
transporte activo:
|
transporte activo ligado a
simporte de protones;
| |
|
transporte
activo ligado a simporte de iones Na+
| |
|
transporte activo dirigido por ATP
| |
|
transporte
acoplado a translocación de grupos.
|
2.3.1 TRANSPORTE ACTIVO LIGADO A SIMPORTE DE PROTONES
Como se recordará, el simporte se puede definir como el transporte
simultáneo de dos sustratos en la misma dirección, por un mismo transportador
sencillo. En el caso del transporte activo ligado a simporte de protones, lo que
ocurre es que uno de los sustratos (H+) ha creado previamente un
gradiente de concentración, cuya disipación es aprovechada por el otro
sustrato para entrar con él. Este otro sustrato puede ser:
|
una molécula de carga negativa:
en este caso, su simporte ligado a protones tiende a disipar sólo el
gradiente de concentración. Ejemplos: transporte de iones fosfato, de
glutamato, etc.
| |
|
una molécula neutra: en este
caso, su simporte tiende a disipar no sólo el gradiente de concentración,
sino también el gradiente eléctrico. Ejemplo: en Escherichia
coli, la lactosa usa una ß-galactósido-permeasa, que es una de las
permeasas bacterianas más intensamente estudiadas.
|
Por
otro lado, ciertas moléculas catiónicas (iones K+, lisina), son
transportadas directamente a través de permeasas, en ausencia de simporte de
protones.
2.3.2 TRANSPORTE ACTIVO LIGADO A SIMPORTE DE IONES SODIO
Se puede considerar una versión modificada del anterior: algunas
sustancias no son transportadas activamente de forma directa por el potencial
electroquímico de protones, sino indirectamente, a través de un gradiente de
Na+ que a su vez se origina a expensas de dicha fuerza protón-motriz
(fpm).
El sustrato entra por una permeasa, junto con iones Na+, pero
a su vez este sodio se recicla por un sistema de antiporte, a expensas de la
disipación del potencial de protones.
Ejemplo: el azúcar melibiosa, en el caso de la enterobacteria E. coli.
Estos dos tipos de transporte activo
ligados a simporte quedan inhibidos si tratamos las células con algún agente ionóforo
(p. ej., el antibiótico valinomicina), que destruye el potencial electroquímico
de protones.
El transporte activo ligado a simporte de iones (H+, Na+)
resulta muy económico, ya que sólo se gasta un protón por cada molécula
transportada, mientras que por cada ATP sintetizado se suelen gastar 3 protones
que se disipan en las ATPasas. Las permeasas que realizan este transporte suelen
ser proteínas integrales de membrana provistas de unos 12 segmentos
transmembranosos en configuración de a-hélice.
2.3.3 TRANSPORTE ACTIVO DIRIGIDO POR ATP
El tipo paradigmático de este tipo de transporte se denomina de transportadores
ABC o ATPasas de tráfico, y
se conocen muchos ejemplos en eubacterias y arqueas. Vamos a describir el
caso de un sistema ABC en enterobacterias (como E. coli). Se trata de un
sistema de varios componentes, en el que existen proteínas periplásmicas que
captan el sustrato con gran afinidad, y lo llevan hasta unas proteínas de
membrana, las cuales acoplan el paso de dicho sustrato hasta el citoplasma (sin
alteralo químicamente) con la hidrólisis de ATP.
La denominación de "transportadores ABC" se debe a que en todos ellos existe una o dos proteínas periféricas de membrana citoplásmica que poseen un dominio (de unos 200 aminoácidos) conservado evolutivamente, denominado "cassette de unión a ATP" (las iniciales de ATP-binding cassette generan la sigla "ABC"). Al parecer este dominio ABC conservado es muy antiguo, y parece que ya existía antes de la divergencia evolutiva entre procariotas y eucariotas. Existen muchos ejemplos de proteínas ABC, tanto en procariotas como eucariotas (de hecho constituyen la mayor familia de proteínas filogenéticamente relacionadas de todo el mundo vivo). Los primeros ejemplos de esta gran familia estaban implicados en transportar sustancias a uno u otro lado de la membrana. En otro tema veremos ejemplos de transportadores ABC que funcionan "al revés" de los de este tema: son "exportadores" de proteínas recién sintetizadas que deben insertarse en las envueltas bacterianas o ser excretadas al medio. Pero se ha descubierto que la gran familia de proteínas ABC está implicada en otros procesos (regulación genética, reparación de ADN, patogenicidad, etc).
Elementos
de este tipo de sistema:
|
Porinas u otras proteínas de
membrana externa para lograr la difusión del sustrato desde el medio hasta
el espacio periplásmico.
| |
|
Proteína(s) solubles de espacio
periplásmico que se unen al sustrato con gran afinidad.
| |
|
Un
heterodímero formado por dos proteínas integrales de membrana (cada una de
ellas posee 5 o 6 trechos en a-hélice
que atraviesan la membrana citoplásmica), que son la permeasa propiamente
dicha del sistema (el canal por donde pasa el sustrato).
| |
|
Dos proteínas periféricas de
membrana citoplásmica, adosadas al lado citoplásmico, que incluyen el módulo
conservado ABC que acopla la hidrólisis de ATP con el transporte
unidireccional del sustrato a través de la membrana.
|
Modelo
del mecanismo de este sistema:
1.
El sustrato exógeno normalmente entra al periplasma a través de algún
canal inespecífico o porina de membrana externa (por ejemplo, en
enterobacterias se pueden usar las porinas generales conocidas como OmpF y
OmpC).
2.
La proteína periplásmica específica, antes de su unión al sustrato
tiene una determinada configuración (denominada “abierta”), con dos grandes
lóbulos globulares unidos formando un ángulo (la forma recuerda una almeja a
medio abrir). Cuando el sustrato pasa al periplasma, la correspondiente proteína
de unión periplásmica se une a él con gran afinidad (0.1-1 mM),
y al unirse cambia de conformación. En esta configuración, llamada
“cerrada”, el sustrato se encuentra “enterrado” entre los dos lóbulos
de la proteína (“almeja cerrada”).
3.
Mientras tanto, el dímero de proteínas integrales de membrana (antes de
la unión con la proteína periplásmica) se encuentra en un estado energizado
pero incapaz de transportar sustrato. En esta situación, puede unirse (por la
parte que da al periplasma) al complejo formado por la proteína periplásmica
(en configuración “cerrada”) ligada al sustrato. Al hacer esto, el heterodímero
de membrana cambia de conformación, de modo que ahora muestra mayor afinidad
hacia la proteína periplásmica y se abre su canal para dejar entrar el
sustrato.
4.
Entonces, el complejo de membrana alcanza su estado de mínima energía,
y con ello descarga el sustrato en el citoplasma y se logra la separación de la
proteína periplásmica (que vuelve a su configuración “abierta”).
5.
Finalmente, la hidrólisis de ATP catalizada por las proteínas periféricas
ABC (adosadas a la membrana y asociadas a las proteínas integrales) suministra
la energía para que el heterodímero de membrana vuelva a su estado energizado
inicial, preparado así para otro ciclo de transporte.
El
sistema ABC de bacterias Gram-negativas, queda puesto de manifiesto cuando lo
impedimos por algún procedimiento que altere o elimine la membrana externa
(conversión a esferoplastos): Por ejemplo: tratemos E.
coli con el quelante EDTA en una solución tamponada isotónica (con un 20%
de sacarosa). Centrifugamos y el sedimento de células lo resuspendemos en una
solución de MgCl2 en frío (0ºC). El resultado es que las proteínas
del periplasma escapan al medio externo. Esto es un caso de tratamiento por choque
osmótico que origina pérdida de contenidos del periplasma. Se comprueba
que ciertos sistemas de transporte quedan inutilizados, debido a que requieren
para su funcionamiento, amén de proteínas
de membrana citoplásmica, otras específicas del espacio periplásmico. Por
contra, este sistema no se ve afectado por los agentes ionóforos. Por esta razón,
a este sistema en Gram-negativas se le ha llamado durante mucho tiempo “transporte
activo sensible a choque osmótico”.
Los sistemas ABC de bacterias
Gram-positivas están menos estudiados, pero en general se parecen a los de
Gram-negativas, salvo que carecen del transportador libre periplásmico. En su
lugar existe una proteína con funciones equivalentes (captar el nutriente del
exterior), pero que está anclada al lado externo de la membrana citoplásmica,
cerca del heterodímero integral de membrana (la unión es mediante su cisteína
N-terminal, que se une a un fosfolípido).
Existen muchos ejemplos de transportadores procarióticos de tipo ABC, y
cada uno de ellos está especializado en transportar un sustrato específico o
varios sustratos parecidos. Ejemplo de sustratos transportados de esta forma:
|
Monosacáridos
como arabinosa, galactosa, maltosa, ribosa, xilosa, etc.
| |
|
Oligosacáridos
| |
|
Iones
orgánicos e inorgánicos
| |
|
Aminoácidos
como histidina, glicina, leucina, etc.
| |
|
Oligopéptidos
| |
|
Algunas vitaminas y metales.
| |
|
Sideróforos
con hierro
|
2.3.4 TRANSPORTE ACOPLADO A TRANSLOCACION DE GRUPOS
Es un sistema de transporte que acopla la entrada del sustrato con su
modificación química por unión covalente con un grupo químico. Estrictamente
hablando, no es un transporte activo, porque no funciona en contra de un
gradiente de concentración, pero se considera de hecho como activo, ya que la
concentración del sustrato modificado
dentro de la célula supera con creces a la del sustrato sin
modificar en el exterior.
Este sistema supone un ahorro de energía metabólica: aunque en el
transporte se gasta un enlace rico en energía, el sustrato queda modificado en
su paso a través de la membrana en la forma que la bacteria emplea como primer
intermediario de su ruta metabólica. Es decir, con un solo proceso se cumplen
dos funciones distintas: transporte y preparación química para la ruta, que de
todas formas habría que realizar. No es de extrañar que este tipo de
transporte haya sido seleccionado frecuentemente en la evolución bacteriana, y
que hoy lo encontremos en muchos procariotas, especialmente en bacterias
anaerobias o aerobias facultativas que recurren a fermentaciones (recordar que
las fermentaciones tienen un rendimiento energético menor que los procesos
respiratorios; por lo tanto, es “lógico” que se seleccionen mecanismos
ahorradores como el descrito).
El caso mejor estudiado de esta clase de transporte lo constituye el
llamado sistema de
fosfotransferasa de azúcares (según las casi inevitables
iniciales inglesas: PTS).
En E. coli el sistema PTS
permite el transporte de glucosa, manosa, fructosa y los polioles sorbitol y
manitol.
Consta de varios componentes que funcionan como una cadena de
transportadores del grupo fosfato de alta energía del fosfoenolpirúvico (PEP)
hasta el azúcar a transportar en cuestión.
|
Las dos primeras proteínas son
inespecíficas respecto del azúcar (son comunes
a los diversos sustratos a transportar), tienen localización
citoplásmica y su síntesis es
constitutiva. Se conocen como
| |||||||
|
El otro componente, llamado Enzima
II (EII) es específico de cada azúcar,
y su síntesis es inducible por el
correspondiente sustrato: Suele estar compuesto por tres subunidades o
dominios:
|
Veamos cómo funciona el
sistema:
1.
Por un lado, el azúcar se une al enzima EIIC específico, pero éste por
sí mismo no puede liberar al azúcar sin modificar en el interior celular.
2.
Mientras tanto, la EI cataliza (en presencia de Mg++) la
transferencia del fosfato de alta energía del PEP a la HPr.
3.
La HPr fosforilada (HPr-P) transfiere el fosfato al enzima IIA específico
del azúcar [p. ej., la glucosa (EIIAGlc ) o el manitol (EIIAMtl)].
4.
La EIIA-P rápidamente, y en presencia de Mg++, transfiere el
fosfato a la enzima-IIB específica con la que se asocia (p. ej., EIIBGlc),
que a su vez fosforila el azúcar (en el caso de la glucosa convirtiéndola en
glucosa-6-P): en este momento la EIIC pierde su afinidad por el azúcar
modificado, que de esta forma entra en el citoplasma, preparado ya para actuar
como sustrato de la primera reacción del catabolismo de este azúcar.
Otros ejemplos de transporte acoplado
a translocación de grupos:
|
Entrada de ácidos grasos mediante
un sistema de transferencia de Coenzima A, que los transforma en acil-CoA.
| |
|
Entrada
de purinas y pirimidinas, mediante un sistema de fosforribosil-transferasas:
|
purina o pirimidina (exterior) + PRPP à
NMP (interior) + P
(PRPP = fosforribosil-pirofosfato)
(NMP = nucleósido monofosfato)
En bacterias es frecuente encontrar varios sistemas de transporte para un
mismo nutriente (p. ej., Escherichia coli
posee cinco sistemas para transportar la galactosa y tres sistemas para algunos
de los aminoácidos. Los diversos sistemas se diferencian en cuanto a su
requerimiento energético, su afinidad, su regulación, etc. Lógicamente, la
evolución ha debido seleccionar esta redundancia de sistemas de transporte con
objeto de permitir que el microorganismo sobreviva bajo diversas circunstancias
ambientales.
2.4 TRANSPORTE DE HIERRO
El hierro es un cofactor de muchas enzimas y citocromos, por lo que las
bacterias necesitan captarlo. La captación de hierro se complica porque el ión
férrico (Fe3+) es muy insoluble. Además, las bacterias que viven
dentro de animales superiores tienen un problema: en los fluidos y tejidos de
sus patrones el hierro libre es muy poco abundante (el hierro suele estar
acomplejado con proteínas), por lo que se vuelve vital aprovisionarse con este
elemento de alguna manera.
Muchas bacterias secretan unas moléculas de bajo peso molecular llamadas
en general sideróforos,
que son capaces de formar quelatos (complejos) con el hierro férrico. Por
ejemplo, Escherichia coli secreta un sideróforo llamado enterobactina.
Cuando la enterobactina se une al hierro, forma un complejo octaédrico, que
luego se engarza con un receptor específico de la membrana externa, tras lo que
el hierro se libera al espacio periplásmico, desde donde entra al citoplasma
por medio de una proteína de unión periplásmica acoplada a un sistema ABC
similar al visto más arriba.
Suscribirse a:
Comentarios (Atom)